New Human Physiology | Paulev-Zubieta 2nd Edition
Chapter 4:Brain Function, Locomotion and Disorders
| HOME | PREFACE | TABLE OF CONTENTS | SYMBOLS | SECTION INFO | CONTRIBUTORS | LINKS | CONTACT US |
Highlights
Study_ObjectivesPrinciplesDefinitionsEssentials
PathophysiologyEquationsSelf-AssessmentAnswers
Further Reading
|
Chapter 4
|
 |
|
The functional unit of the nervous system is the neurone with its cell body, dendrites and axon, which terminates in a synapse. Action potentials passing down the axon release chemical neurotransmitters at the synapse.
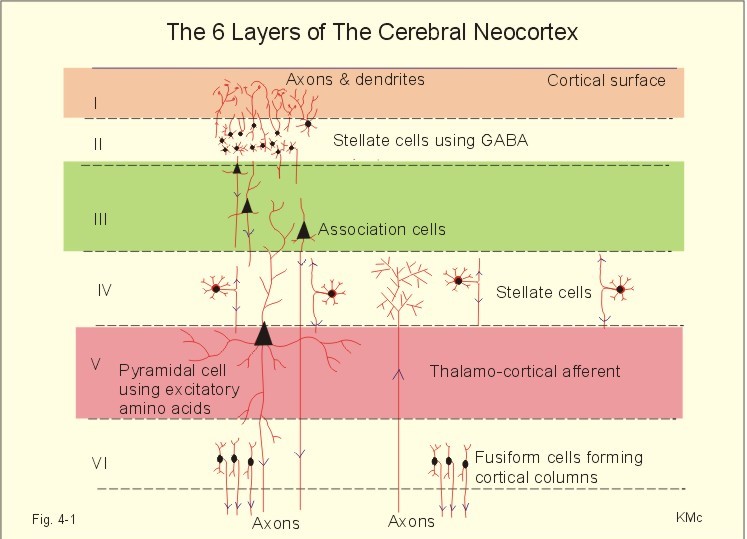
This paragraph deals with 1. The cortex and the reticular activating system, 2. Higher brain functions, 3. The limbic system and the hippocampus, 4. Spinal organization of the motor control, 5. Descending motor pathways, 6. Motor control by the brain. 1. The Cortex and reticular activating system Six cell layers are recognized in typical regions of the cerebral neocortex and they are numbered layers I-VI (Fig. 4-1). The neocortex first appears with the mammals, and its structure is phylogenetically younger than the allocortex. All six layers contain glial cells and more or less of the three typical neurons: Stellate cells, pyramidal cells and fusiform cells. The superficial layers receive and process information, whereas the deep layers are the sites of origin of most cortical efferents. The six neocortical layers are as follows:
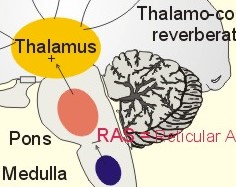
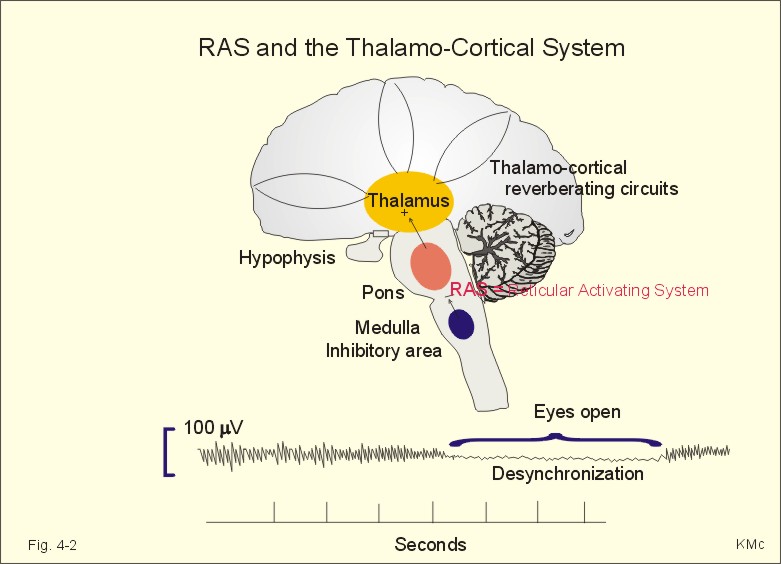
Fig. 4-1: The cerebral neocortex with pyramidal, stellate and fusiform cells. The pyramidal and fusiform cells of layers V and VI provide the output from the cortex. The pyramidal cells have long axons passing to other cortical regions to the brain stem and to the spinal cord. Thalamocortical afferents mainly project to layers I, IV, and VI, whereas corticothalamic projections have their origin in pyramidal cells in layers V and VI. These connections form a reverberating thalamocortical system, which excite the cortex and contribute to the patterns of the electroencephalogram (EEG). A large region of the reticular formation of the brainstem is termed the ascending reticular activating system (RAS), which determines our state of consciousness, by its connection with the thalamocortical system (Fig. 4-2). The RAS transmits facilitatory signals to the thalamus. The thalamus excites specific regions of the cortex, and the cortex then excites the thalamus in a reverberating circuit with fast acetylcholine and long-lasting neuropeptides as transmitters. Such a positive feedback loop is what wakes us up in the morning. External stimuli and internal factors (inhibitory interneurons) serve to create a balance of different activity levels during the day. RAS maintain the ascending thalamic activity, but also a certain descending activity level in our antigravity muscles and reflexes. An inhibitory region in the medulla can inhibit RAS and thus both its ascending and descending activity. A high level of consciousness is called arousal or alertness, which is recognized in the EEG as a high frequency-low voltage shift (see below). An orientation reflex, a fundamental change of behaviour following an external stimulus, often accompanies arousal. The eyes, head and body are turned toward the external stimulus. Impaired consciousness is caused by malfunction of the neurons in the RAS, and the impairment has at least three levels. Sopor or clouding of consciousness is a term for reduced wakefulness, stupor is a sleepy state from which the patient can be aroused by vigorous stimuli, and coma is a unresponsive state of unconsciousness from which the patient cannot be awakened even with the most vigorous stimuli. Brain stem compression at the mesencephalon leads to coma and death.
Fig. 4-2: The RAS and the thalamocortical system. The electroencephalogram (EEG) is a rhythmic electrical activity recorded from the surface of the skull. In humans, the EEG is recorded from a grid of standard leads (Fig. 4-2). During neurosurgery the electrical activity is recorded from the surface of the cortex as an electrocorticogram. In normal adult persons, the dominating frequencies are 8-13 Hz (a-rhythm) over the parietal and occipital lobes, as long as the subject is awake and relaxed with his eyes closed. With open eyes, the EEG becomes desynchronized with low amplitude (10 mV) and the dominant frequency increases to 50 Hz. The theta- (3-7 Hz) and delta rhythms (0.5-2 Hz) are observed during light and deep sleep, respectively. A thalamocortical rhythm produces coordinated extracellular currents, when the brain is not exposed to external stimuli (Fig. 4-2). The EEG recording is due to large synaptic potentials by whole groups of mainly pyramidal cells. The EEG pattern is desynchronized by sensory inputs through the thalamus. The level of alertness (in RAS) also modifies the EEG pattern. Each pyramidal cell - as with each Purkinje cell in the cerebellum - posses an extraordinarily large number of synapses (106). The potentials recorded on the surface of the skull are 50-100 mV. The large pyramidal cells form a dipole with one pole directed toward the surface of the cortex, and the other toward the white matter. When an external stimulus evokes an EEG change, the change is termed a cortical evoked potential. The large numbers of synaptic potentials in the cortical region are summated to form an evoked potential recorded on the skull by an electrode placed over the associated cortical area. However, evoked potentials are small, so measurement requires repeated stimulation and signal averaging. The evoked potentials over the auditory, visual, and somatosensory cortex (areas I and II) are used clinically to assess the integrity of the respective sensory pathway. Circadian periodicities are changes in biological variables that occur daily. The circadian controller is the so-called biological clock, probably located in the suprachiasmatic nucleus of the hypothalamus. The biological clock receives many projections from sense organs including projections from the retina signaling light and darkness. These signals are transmitted further to the pineal gland according to one hypothesis. Darkness probably stimulates melatonin secretion by the pineal gland, which inhibits the secretion of gonadotropic hormones from the anterior pituitary, and thus reduces sexual drive. Melatonin secretion decreases with age. Destruction of the biological clock disrupts many biological rhythms, such as oscillations in body temperature, other vegetative functions and the sleep-wake cycle. The astronomic 24-hour cycle is shorter than the biological sleep-wake cycle (normally 25 hours). When flying east the astronomic cycle is shortened further acerbating the discrepancy between the two cycles. This increases the problems of adjusting the circadian systems, which often require a week to regain their normal phase relation to the biological clock. Problems caused by changes of biological rhythm are summarized in the term jet lag. Melatonin is used clinically to reduce the jet lag. The endogenous circadian periodicity of the sleep-wake cycle is normally 25 hours - see above. Sleep is divided into four stages based on EEG. The relaxed individual with eyes closed has 8-13 Hz a-rhythm. As he falls asleep, he passes through the four stages of sleep. During these stages the muscles are relaxed, all vital functions are decreased, and the gastrointestinal motility is increased. Stage 1 is light sleep, where a-rhythm is interspersed with theta rhythm. Stage 2 is somewhat deeper sleep dominated by slow waves and by sleep spindles (periodic spindle-shaped bursts of a-rhythm) and by large, irregular K-complexes. Stage 3 is characterized by delta waves and by occasional sleep spindles. Stage 4 is recognized by the very slow delta waves with frequencies around 0.5-1 Hz. The subject is difficult to wake up. A different form of sleep with complete loss of muscle tone occurs periodically every 90-min during stage 1 sleep. This is termed rapid eye movement sleep or REM sleep. Eye movement artifacts and a desynchronized EEG (low voltage, fast activity as in the arousal reaction when awake), is characteristic for REM sleep. The subject is difficult to wake up, so the condition is therefore also termed paradoxical sleep.
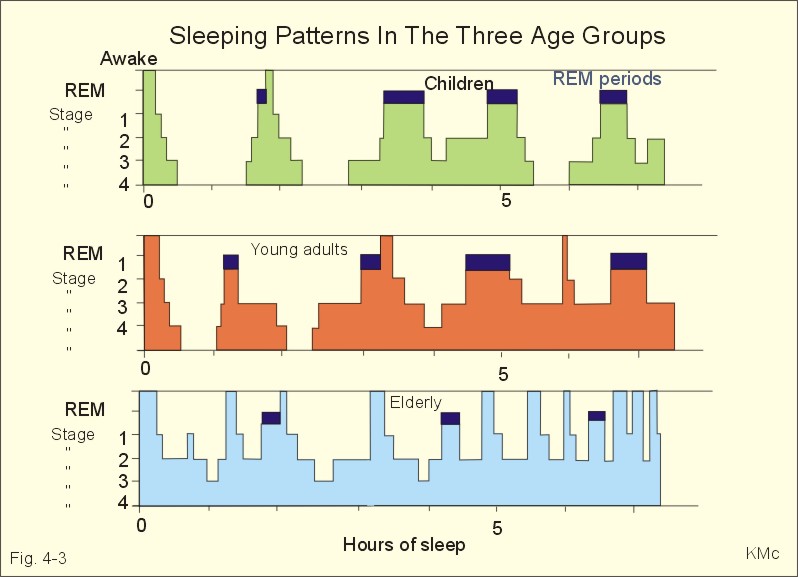
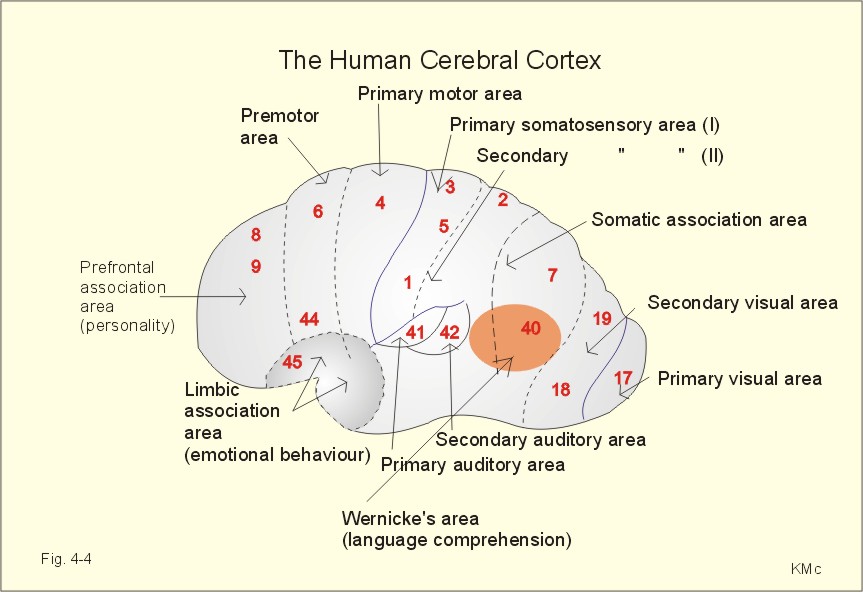
Fig. 4-3. Differences in sleeping pattern between three age groups. Spontaneous erection occurs during REM sleep, and an irregular heart rate and respiration are often observed. Dreams occurring during REM sleep are often recalled by the person when awake. Children and young adults have all 4 stages of sleep and several periods of REM sleep (Fig. 4-3). The depth of the non-REM sleep diminishes through the night and the REM periods increase in duration (Fig. 4-3). Stage 4 sleep disappears with age, and stage 3 sleep decreases in duration (Fig. 4-3). The REM sleep is also reduced, and wake periods occur in increasing number. This is why elderly people believe that they do not sleep sufficiently. The passive theory of sleep claim sleep to be caused by reduced activity in RAS. However, transecting the brainstem in the midpontile region produces an animal that never goes to sleep. Stimulation at the nucleus of the solitary tract can induce sleep, suggesting that sleep be an active process related to centres below the midpontile level. The question is difficult to address. An educated guess is that sleep is an active, energy saving condition, preferable to most animals. The metabolic rate during sleep falls to 75% of the basal metabolic rate. Each hemisphere consists of the following four lobes: the frontal - occipital - parietal and temporal lobes. The frontal lobe, located in front of sulcus centralis (central fissure), is involved in motor behaviour. The frontal lobe contains the primary motor (area 4), the premotor (area 6), and the supplementary motor areas (frontal eye areas 8 and 9 of Fig. 4-4). These cortex areas are responsible for planning and execution of voluntary movements. The motor speech areas (44 and 45 or Broca's area) are located close to the motor cortex, on the inferior frontal gyrus of the dominant hemisphere in humans (the left hemisphere is controlling the expressed language in most people). Lesions here cause motor aphasia (difficulties with speech and writing). Patients with lesion of Broca's area (in the dominant hemisphere) frequently suffer from paralysis of the opposite side (right) of the body.
Fig. 4-4. The human cerebral cortex of the left hemisphere controls the expressed language. The frontal cortex is also involved in personality and emotional behaviour - including attention, intellectual and social behaviour. The occipital lobe is located behind the parietal and temporal lobe, and involved in visual processing and visual perception. Adjustments for near vision are controlled by the primary visual cortex in area 17 and in the cortex around the calcarine fissure occipital lobe. The conscious visual perception takes place in the primary visual cortex. The secondary visual cortex is in area 18 and 19, where visual impressions are compared, interpreted and stored (Fig. 4-4). The important primary somatosensory area I is located on the postcentral gyrus (area 1,2 and 3 in Fig. 4-4). There is a distinct spatial representation of the different areas of the body in the postcentral gyrus (the sensory homunculus). The secondary somatosensory area II is located in the rostral part of area 40, close to the postcentral gyrus (Fig. 4-2). The somatic association or interpretation areas (areas 5 and 7) are located in the parietal cortex just behind the somatosensory area I (Fig. 4-2). Each side of the cortex receives information exclusively from the opposite side of the body. Widespread damage to the somatosensory area I causes loss of sensory judgement including the shapes of objects (astereognosis). Auditory and vestibular signals are processed and perceived by the superior temporal gyrus (area 41 in Fig. 4-2). Area 42 is the secondary auditory centre, where auditive signals are interpreted and stored. The medial temporal gyrus helps control emotional behavior in the limbic system and all the functions of the autonomic nervous system. Signals from the auditive (area 42), visual (areas 18 and 19) and somatic (areas 7 and 40) interpretative areas are integrated in the posterior part of the superior temporal gyrus. This large gnostic area is specially developed in the dominant hemisphere, where it is called the general interpretative or language comprehension area (Wernicke's area). Damage in Wernicke's area causes sensory aphasia (i.e., difficulties in understanding written or spoken language, although single words can be heard). Learning is a change of behavior caused by neural mechanisms affected by experience. Memory refers to neural storage mechanisms for experiences. The hippocampus is involved in learning and memory.
Habituation refers to a gradual diminution of a response by repetition of a stimulus, because experience show that the stimulus is unimportant. Sensitization is the opposite of habituation. Firstly, a strong threatening stimulus triggers a certain response, but repetitions of the stimulus increase the size of the response in order to avoid the threat. This evaluation is called the reward and punishment hypothesis. The neural processes are probably related to the function of the hippocampus. In the snail aplysia a facilitating interneuron releases serotonin onto the presynaptic terminal of a neuron. This stimulates adenylcyclase and the formation of intracellular cAMP in the presynaptic terminal. The resulting protein kinase activation causes phosphorylation and blockage of K+-outflux. The K+-outflux is necessary for recovery from the action potential. Lack of K+-outflux prolongs the presynaptic action potential considerably. This causes a prolonged Ca2+-influx into the presynaptic terminal with increased release of neurotransmitter and facilitated synaptic transmission.
Conditioning refers to a neural process of associative learning, where there is a temporal association (optimum 0.5 s) between a neutral stimulus (eg, a sound before food) and an unconditioned stimulus (food) that elicits a response (gastro-intestinal secretion). Repetition of the sound-food manoeuvre develops into a conditioned reflex, where the sound alone elicits salivary secretion. In operant conditioning the response is associated with reinforcement, which changes the probability of the response. Positive and negative reinforcement increases the probability of the response, whereas punishment reduces its probability. Learning is highly improved by happiness. Light stress is an advantage in learning something new. However, substantial stress is not helpfull in the recall process, and stress can completely block the memory. Strategic behavior is the basis for our social life. Strategic or motivated behavior is related to homeostasis in general (defence, reproduction, temperature and appetite control). Previously, strategic behavior was explained by negative feedback with the purpose as a fixpoint, and with the human brain playing a minor role. Today it is generally accepted that the cerebral drive is a dominant determinant for strategic or motivated behavior. The drive that arouses individuals from inactivity originates in the limbic system (including the hypothalamus), that is acting in close relation to the thalamus and the cerebral cortex. The limbic system is connected to the autonomic control functions of the brainstem reticular formation by the medial forebrain bundle. These vital functions are thermocontrol, appetite control and sexual behavior (Chapter 6). The dominant hemisphere is the hemisphere that controls the expressed language. Lesions of the left hemisphere produce deficits in language function of most people. These deficits are called aphasia. The left planum temporale in the floor of the lateral fissure of Sylvii is larger than that of the right hemisphere in most people - not only right-handed. The right hemisphere is dominant for functions related to language (intonation, body language), and to mathematically related functions. Each hemisphere controls the contralateral side of the body. Information between the two hemispheres is transferred through the anterior commissure and the corpus callosum. The language centres on the left hemisphere cannot influence the right hemisphere unless the corpus callosum is intact. The two hemispheres can operate relatively independently with language. One hemisphere can express itself through spoken language. The other communicates non-verbally. If an animal with intact corpus callosum and optic chiasm learns a visual discrimination task with one eye closed, the task can still be performed with the untrained eye alone, even when the optic chiasm is transected before the animal is trained. Therefore, visual information is transferred as long as the corpus callosum is intact. Surgical transection of the corpus callosum has been performed to prevent epilepsy from spreading. When such a patient fixate his vision on a point on a screen, it is possible to stimulate only one hemisphere by showing an object to one side of the visual field. Similar objects (key, ring, nail, fork etc) can be manipulated (but not seen) through an opening below the screen. Healthy persons can locate the correct object with either hand. Split-brain patients, with the picture of the object transferred to the right hemisphere, can locate the correct object with the left hand (ie, right hemisphere), not with their otherwise preferred right hand. Jigsaw puzzles are solved with such manipulo-spatial capabilities. Right-handed patients with split brain can solve three-dimensional puzzles, if the visual signals can reach the motor cortex for the hand to explore. The visual and motor cortex are connected to each other only in the same hemisphere, when the corpus callosum is cut. Memory research has characterized three temporal stages in human memory processes. 1. An immediate memory holds sensory information for a few hundred milliseconds to seconds for analysis and further processing. The immediate memory is erased by new incoming signals, so we can only remember a few new telephone numbers at a time. Accumulation of Ca2+ in the presynaptic terminals with each signal possibly causes prolonged release of neurotransmitter at the synapse (synaptic potentiation). 2. The short-term memory is covering seconds to a few minutes, and the short-term memory receives selected information from the immediate memory. Information is erased as new items displace old data. If a person sees a rapid succession of slides, it is the last slide that remains in the short-term memory. We store recent events in the short-term memory, by a neural activity with improved synaptic efficacy that lasts for seconds to minutes. The improved synaptic efficacy is possibly due to synaptic potentiation, presynaptic facilitation, or impulses circulating in neuronal circuits for a restricted period. 3. The long-term memory is a large and permanent memory. The long-term memory receives information from the immediate and the short-term memory. Recycling of information through the short-term memory is termed rehearsal. The likelihood of a successful storage in the long-term memory increases with the number of cycles. When the long-term memory is searched for a certain information, it may take minutes to recall the memory. The long-term memory is subdivided into the intermediate long-term memory, which lasts for days or weeks and can be disrupted, and the long lasting long-term memory, which lasts for years. 4 The long lasting long-term memory is the storage in the brain of highly overlearned information as one's own name and address. This memory is difficult to disrupt, and it is seldomly affected in retrograde amnesia (see below). The long-term memory and consolidation of memory relate to effector protein synthesis at the synapses. Electron microscopy suggests an increased number of vesicular release sites in the presynaptic terminals. Retrograde amnesia is a term used for a condition where the patient cannot recall information from the immediate and short-term memory. The mild form of retrograde amnesia is typical following head lesion with loss of consciousness (cerebral commotion). The short-term memories have only been rehearsed a few times and probably stored only discretely. The long term-memories are widespread in the cortex as structurally maintained modifications of the synapses after many rehearsals. Only in severe cases is the long-term memory involved. 3. The limbic system, the hippocampus and emotions The limbic system is the neuronal network that controls emotional and motivational behavior. Motivational behavior include control of vegetative functions such as body temperature, respiration, circulation, osmolality of body fluids, sexual behavior, smell, thirst, appetite and body weight. Hypothalamus constitutes the major part of the limbic system, and is located in the middle of the other limbic elements.
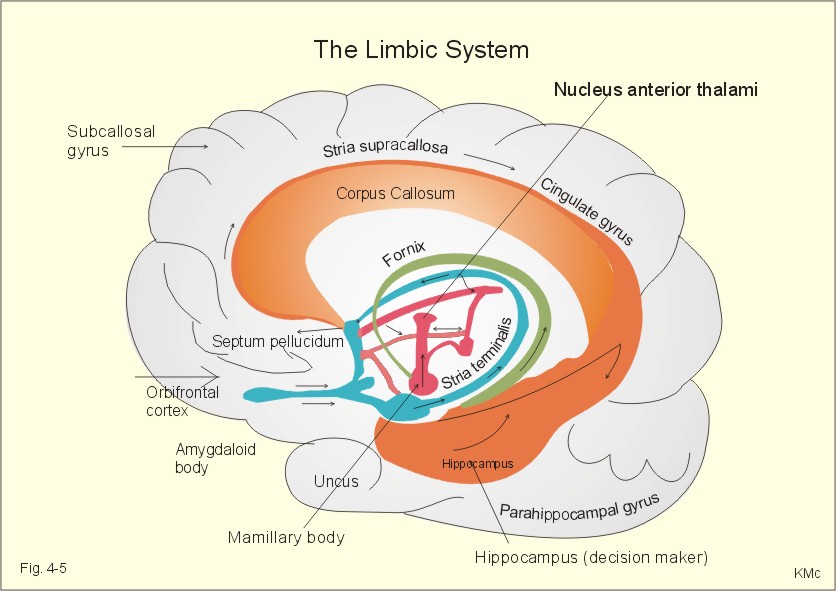
Fig. 4-5. The limbic system. The corpus callosum is transected, and we are looking at the medial aspects of the right hemisphere. The limbic cortex begins in the frontal lobe as the orbitofrontal cortex, extends upward as the subcallosal gyrus, over the corpus callosum and into the cingulate gyrus (Fig. 4-5). The limbic cortex finally passes caudal to the corpus callosum down towards the hippocampus, para-hippocampal gyrus and uncus at the medial surface of the temporal lobe (Fig. 4-5). The fornix connects the hippocampus to the mamillary body. The mamillothalamic tract connects the mamillary body to the anterior nucleus of the thalamus. Thalamus connects to the cingulate gyrus, and its cortex is associated with the hippocampus. Stria terminalis connects the amygdaloid body to the midbrain septum and to the mamillary body (Fig. 4-5). The limbic paleocortex links the subcortical limbic structures to the neocortex. Hereby, the limbic system relates behavior and emotions to the intellectual cortex functions. Another important pathway is the medial forbrain bundle, which connects the limbic system to the autonomic control functions of the brainstem reticular formation. The hippocampus connects with the cerebral cortex, the midbrain septum, the hypothalamus, the amygdaloid and the mamillary bodies and acts both as a store and a recall centre (Fig. 4-5). The hippocampus is the decision-maker, determining the importance of incoming signals. Hippocampus becomes habituated to indifferent signals, but learns from signals that cause either reward (pleasure) or punishment. Hippocampus is the "brain librarian" (helps the cortex to store new signals into the long lasting long-term memory). The signal molecule, nitric oxide (NO), modulates aspartate responses related to hippocampal long-term potentiation. Bilateral removal of the hippocampi in epileptic patients permanently disrupts the ability to learn anything new (anterograde amnesia). Other lesions of the hippocampi reduce previously learned memory material (retrograde amnesia - see above). Long-term alterations imply a rise in the number of synapses. Cholinergic synapses in the midbrain septum are essential to our memory, and these neurons are dependent upon the nervous growth factor. Repeated activation of a sensory pathway increases the reaction of pyramidal cells. Such a reaction may last for weeks in the hippocampus and be involved in storage and retrieval of new information in the long-term memory. Our memory (cortex and hippocampus) works as a filter. Perhaps only 1 per mille of all received signals contain useful or emotional information and are catched in the memory. Unfortunately, we are unreliable witnesses, because we invent emotional "information" concerning a factual experience. The easiest facts to remember are those that make sense. All facts, concepts and acquired skills are stored in a ready-to-use fashion. Feelings play a large role in memory, and strong impressions that are charged with emotion etch themselves into our memory. A recollection is split up into numerous subunits in different regions of the brain. Later, all subunits are brought together by the hippocampus into a complete memory (eg, a certain smell act as a strong clue to a clear memory from way back). One individuals recollection of a particular incident can trigger off anothers, whereby new associations can be created. 4. Spinal organization of motor control Motor activity can be voluntary or involuntary. Voluntary movements are planned and started by feedforward control, and when maintained for a while they are regulated by feedback loops. Involuntary movements comprise reflexes, such as the stretch reflexes, and autonomic functions, such as the respiratory muscle movements. We have motor centres in the cerebral cortex, the brainstem, the spinal cord, the cerebellum, and the basal ganglia. Motor centres all receive sensory information in an organized neural structure termed a somatotopic map (see the motor homunculus). We have 200 different skeletal muscles, which are controlled by more than 300 000 motor units. A motor unit is comprised of a a-motor neuron, all its axon terminals, and the skeletal muscle fibres it innervates. The number of muscle fibres in a motor unit varies from 2 in highly regulated eye muscles (entirely red fibres) to 2000 in the quadriceps femoris muscle. The motor unit is the final common pathway, because all muscle fibres of the unit contract when a motor unit is activated. Adjacent motor units interdigitate, so they can support each other. The muscle power is increased by recruitment of more motor units and by increased frequency of discharge in each unit. We have three types of motor units (a-motor neurons) in a mixed muscle such as the gastrocnemius. The three types of motor units are characterized in Chapter 2, Table 2-2. A spinal reflex is a stereotyped motor reaction to an input signal. The myotatic stretch reflex is the most crucial monosynaptic reflex for the maintenance of the erect body posture in humans.
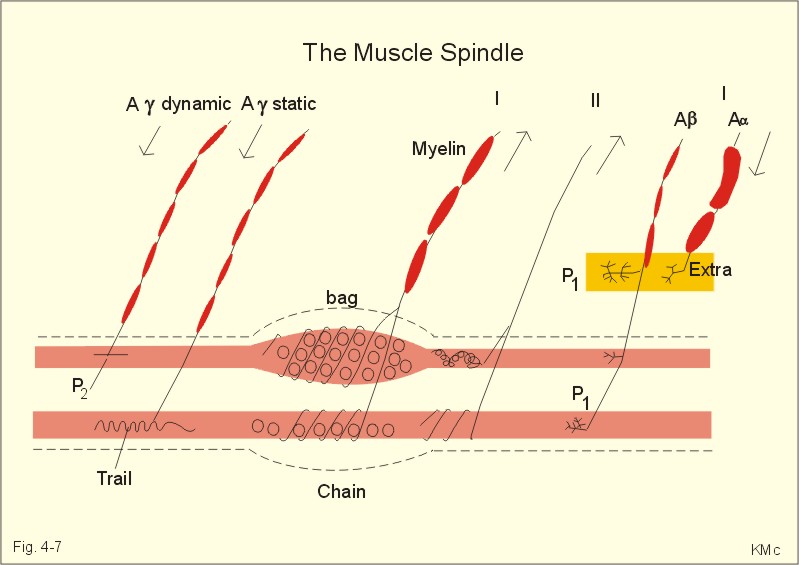
Fig. 4-6: The phasic myotatic stretch reflex and reciprocal innervation (F-). The reflex has two components. Firstly, the primary annulospiral endings (group Ia) of the muscle spindles trigger the phasic stretch reflex. Secondly, both primary and secondary endings elicit the tonic stretch reflex. 1. The phasic stretch reflex is elicited in the clinic by a light tap on a muscle tendon. When the patellar tendon from the quadriceps muscle is stretched quickly by the tap, a discharge is elicited in the afferent fibres (Ia) from the primary endings of the muscle spindle (Fig. 4-6). This is the phasic myotatic stretch reflex or the so-called patellar reflex. These Ia fibres synapse directly (monosynaptically) on a-motor neurons that supply the extensor muscles of the knee (E+ in Fig. 4-6). The response elicited is a brief contraction of the latter. Of all the presynaptic terminals arriving to the motor neuron up to 90% are located on the surface of the dendrites. The remaining 10% synapse on the soma of the motor neuron. The Ia afferent fibres also synapse with small group Ia inhibitory interneurons in the grey matter of the spinal cord, as the one synapsing with the upper a-motor neuron in Fig. 4-6. This neuron innervates the semitendinosus muscle, which flexes the knee joint (F- in Fig. 4-6). The reflex inhibition of antagonist muscles when synergistic muscles are contracted is called reciprocal innervation. In pathologic conditions, the phasic stretch reflexes may be depressed or hyperirritable. 2. Passive bending of a joint triggers the tonic stretch reflex. This elicits a discharge in both groups Ia and II afferents from the muscle spindle. The tonic stretch reflex contributes to the erect body posture and helps maintain posture by increasing the tone of the physiologic extensor muscles (ie, antigravity muscles). Renshaw inhibition and presynaptic inhibition Renshaw inhibition. Cajal found that the a-motor axons give off thin recurrent (antidromal) collaterals in the grey matter of the spinal cord (Fig. 4-6). These collaterals synapse with Renshaw interneurons in the ventral horn (Fig. 4-6). The Renshaw cells synapse with a-motor neurons of synergistic muscles, and thus inhibit monosynaptic reflexes (postsynaptic inhibition). Stimulation of each a-motor unit inhibits adjacent motor units (ie, recurrent inhibition). This is also called the principle of lateral inhibition, whereby the motor response is confined to selected units only. Descending signals from the brain can either amplify the postsynaptic inhibition or reduce its effect. Renshaw cells make it possible for the higher brain centres to influence spinal reflexes by central inhibition or facilitation. Presynaptic inhibition. Presynaptic terminals contain a large number of voltage-gated Ca2+-channels. Ca2+ must enter the presynaptic terminal from the extracellular space before the vesicles can release their neurotransmitter at the synapse. Presynaptic inhibition takes place at presynaptic contact sites on the presynaptic terminals. Activation of these sites closes many Ca2+-channels, and thus inhibits transmitter release. The Golgi tendon organs are the serially located terminals of group Ib fibres wrapped around bundles of collagen fibres in the tendons. Golgi tendon organs monitor the force in the tendon; they are activated either by stretch or by contraction of the muscle. The adequate stimulus is the force developed in the tendon. The inverse stretch reflex or the Golgi tendon reflex completes the stretch reflex by a force-controlling feedback. The Golgi tendon organs monitor force in the tendons. Golgi tendon organs are in series with the muscle fibres - not parallel as the muscle spindles. If the extensor muscles of the thigh are fatigued, as during standing, the force in their tendons begins to decrease. This reduces the discharge of the Golgi tendon organs. This acts as a compensating feedback, which excites the a-motor neurons and increases the force of contraction. The inverse stretch reflex helps maintain the force of muscular contraction and posture during standing. During the rapid contraction of the myotatic stretch reflex, the inverse stretch reflex reduces the force of contraction. The stretch reflexes regulate the length of the muscle, and provide a length-force feedback to the CNS. The muscle spindle monitors muscle length and rate of change of length (velocity); they are particularly abundant in muscles that are capable of fine movements and in large muscles that are dominated by slow twitch fibres. The organ is shaped like a spindle, which lies in parallel to the large, regular, extrafusal muscle fibres. Each organ contains two main types of intrafusal muscle fibres: Nuclear bag fibres which swell in the equatorial region due to all the nuclei located here, and thin nuclear chain fibres which have central nuclei arranged in line (Fig. 4-7). The primary afferent fibres (Ia) twine around the equatorial regions of both the bag and chain fibres like a corkscrew or annulospiral; the annulospiral nerve endings signal length and velocity. The secondary afferent fibres originate mainly from the nuclear chain fibres and with a few branches originating from the nuclear bag fibres (Fig. 4-7). They monitor only the length of the muscle. Two types of g-motor neurons innervate the muscle spindle. The dynamic g-motor axons form plate endings (P2) on the nuclear bag fibres, while static g-motor axons form creeping trail endings on nuclear chain fibres (Fig. 4-7). The intrafusal fibres receive a Ab-motor fibre, which terminates with P1 plate endings on both extra- and intrafusal muscle fibres (Fig. 4-7). The Ab-motor fibres may be involved in a-g-coactivation. When the extrafusal fibres contract, the muscle spindles shorten, whereby the discharge rate of their afferents decreases.
Fig. 4-7: The structure of a muscle spindle with a bag and a chain fibre. Activity of the g-motor neurons causes the polar spindle regions to contract on either end. This elongates the equatorial regions so that muscle spindles can adjust to stretch (Fig. 4-7). Descending commands from the brain often cause contraction of both extrafusal and intrafusal fibres simultaneously so that the muscle spindle is sensitive to stretch at all muscle lengths. When the muscle is stretched, the muscle spindles are simultaneously stretched with it, and the discharge rate of the afferents is increased. The flexion reflexes are triggered by various flexion reflex afferents including nociceptors. The flexion reflexes have a long latency, because it involves polysynaptic interneurons. The afferent discharge causes excitatory interneurons to activate a-motor neurons that innervate ipsilateral flexor muscles. The afferent discharge also causes inhibitory interneurons to inhibit a-motor neurons, supplying the ipsilateral extensor antagonists. The flexor withdrawal reflex is crucial. This reflex is also called a nociceptive reflex or a pain reflex, and involves all the muscles of a limb in flexor withdrawal in order to protect from further damage. In addition, the reflex can activate the extensor muscles of the opposite limb. This contralateral activity is termed the crossed extension reflex by reciprocal innervation. The locomotor pattern generator controls flexion reflexes involved in locomotion. Severe visceral disease can trigger contraction of the chest and abdominal muscles, which reduces pain by limiting movement of the body. When examining the abdomen of such a patient it will be observed that the muscles are tense. This sign is called defence musculaire, which is a viscero-somatic protective reflex. Coordination of limb movements We possess pattern generators or neural circuits in the spinal cord, for every limb and for respiration, chewing etc. The midbrain locomotor centre, via the reticular formation and through the reticulospinal tracts, organizes the commands. Such spinal pattern generators also account for other movement patterns like scratching, dancing etc. Clinical dichotomy traditionally subdivides the descending fibres into the pyramidal and the extrapyramidal pathways; this is based on the fact that the corticospinal tract passes through the medullary pyramids. Therefore, interruption of the corticospinal or pyramidal tract was supposed to cause pyramidal tract disease (see later). The problem, however, is that the loss of the corticospinal tract does not explain all the classical signs of pyramidal tract disease. The concept of extrapyramidal pathways raises other problems. The concept of extrapyramidal tract diseases is generally used to designate one or more disorders of the basal ganglia. While, extrapyramidal pathways do play a role in basal ganglia diseases (as in cerebellar disease), the main motor pathway involved in basal ganglia diseases is the corticospinal tract! The descending motor pathways can also be dichotomized based on their endpoint in the spinal cord, and hence which muscles they control and how. Pathways ending in the lateral horn of the spinal cord (on motor neurons or interneurons) are called the lateral descending motor system (the rubrospinal tract and the lateral corticospinal tract). Pathways ending on the medial ventral horn interneurons are termed the medial descending motor system (containing reticulo-, tecto-, and ventriculo-spinal tracts). The lateral corticospinal, the corticobulbar (to the facial motor and hypoglossal nucleus) and the rubrospinal tracts control the manipulative movements of the limbs and the lower face and tongue muscles. The corticospinal and corticobulbar tracts originate from areas 4, 6, 8, 9, and somatosensory area I (areas 1, 2, 3 in Fig. 4-4). The large and small pyramidal cells and the giant pyramidal cells of Betz are the cells of origin of these tracts. The corticospinal tract descends through the internal capsule and brainstem. At the medullary pyramid 80% of the fibres cross to the opposite side and descend in the dorsal lateral funiculus as the lateral corticospinal tract. The fibres of this tract end on motor neurons and interneurons in the lateral horn of the spinal cord. These motor neurons innervate distal muscle groups. Interruption of the lateral corticospinal tract implies loss of the fine control of the digits. Interruption of the corticobulbar tract to the facial motor and hypoglossal nucleus implies loss of voluntary movements of the lower face and tongue. Interruption of the rubrospinal tract from the red nucleus combined with corticospinal lesions give rise to difficulty in separating finger, hand and arm movements. The red nucleus is closely linked to the deep cerebellar nuclei. The lateral or dorsolateral descending system allows the primary motor cortex to modify the reflexes and pattern movements at the level of the spinal cord. The medial or ventromedial descending system involves the ventral corticospinal tract and much of the corticobulbar tract ending in the medial group of brainstem and spinal cord interneurons. The ventral corticospinal tract continues caudally in the ventral funiculus on the same side and ends bilaterally on the medial interneurons. They control the axial muscles and bilateral activity including chewing and wrinkling of the eyebrows. Other medial system pathways originate in the brainstem: 1. The lateral vestibulospinal tract excites motor neurons that innervate proximal postural muscles. It receives input from all compartments of the vestibular apparatus and from cerebellum to the lateral vestibular nucleus. 2. The medial vestibular tract receives signals from the semicircular ducts and from cerebellum, and excites motor neurons in cervical and thoracic segments. Thus, it controls the head position in response to angular accelerations of the head. 3. The pontine reticulospinal tract excites motor neurons to the proximal extensor muscles to support posture. 4. The medullary reticulospinal tracts have mainly inhibitory effects on many spinal reflexes. 5. The tectospinal tract from the superior colliculus causes contralateral movements of the head in response to touch and auditory stimuli. This tract allows the integration of hearing and vision with motor performance. 6. Pathways from the solitary nucleus and the interstitial nucleus of Cajal are involved in the pharyngeal stage of swallowing. The solitary nucleus receives all sensory signals from the mouth including taste, and is involved in cardiovascular and respiratory control. The ventromedial system is important for the normal muscle tone and body posture. Monoaminergic descending pathways 1. The neurons of the pontine locus coeruleus and nucleus subcoeruleus contain nor-adrenaline (NA). These nuclei project to and inhibit interneurons and motor neurons of the spinal cord through the lateral funiculi. 2. The neurons of the raphe nuclei in the medulla, which are connected to the limbic system also, contain serotonin. The serotonergic nuclei project to and inhibit dorsal horn interneurons reducing pain transmission, and they also project to and excite ventral horn motor neurons of the spinal cord, thereby enhancing motor activity. 3. There is also a descending dopamine pathway. The three monoaminergic pathways function as motor system amplifiers. The primary motor cortex (area 4) on the precentral gyrus controls distal muscles of the extremities. Area 4 is organized parallel to the somatosensory cortex. The face is represented laterally near the Sylvian lateral fissure, and the legs on the medial part of the hemisphere. The cortical representation is somatotopic and disharmonic, as indicated by the motor homunculus. The premotor cortex helps control proximal and axial muscles. The supplementary motor cortex is involved in motor planning and in coordination of movements. The frontal eye fields initiate saccadic eye movements. Corticospinal neurons discharge before voluntary muscle contraction, and the size of the discharge is related to the size of the contractile force. The somatosensory cortex and the posterior parietal association cortex receive feedback from the sensory neurons system, which helps correct motor feed-forward commands. The little brain, also termed the motor autopilot, helps regulate movements and posture, influences muscle tone, eye movements and balance. Cerebellum is particularly concerned about the timing of rapid muscular activities including the interplay between agonist and antagonist muscle groups. Motor learning is programmed in the cerebellum. Cerebellum compares the proprioceptive input from the actual movements, with the movements intended by the motor control areas of the brain. Cerebellum controls the sequence of movements, and makes corrective adjustments just like an autopilot. The cerebellar cortex is characteristically folded and consists of three phylogenetically different structures related to three afferent pathways (inputs). The large neocerebellum in higher mammals is also called the pontocerebellum and consists of the hemispheres and vermis caudal to the primary fissure. The paleocerebellum or spinocerebellum consists of vermis of the anterior lobe, pyramis, uvula and paraflocculus. The small archicerebellum or vestibulocerebellum is simply the flocculonodular lobe. Three important outputs from the cerebellum also divide it into three functional units. The vermis of the cerebellar cortex projects to the fastigial nucleus, the pars intermedia to the globose and emboliform nuclei, and finally the hemisphere, which projects to the large dentate nucleus (Fig. 4-8).
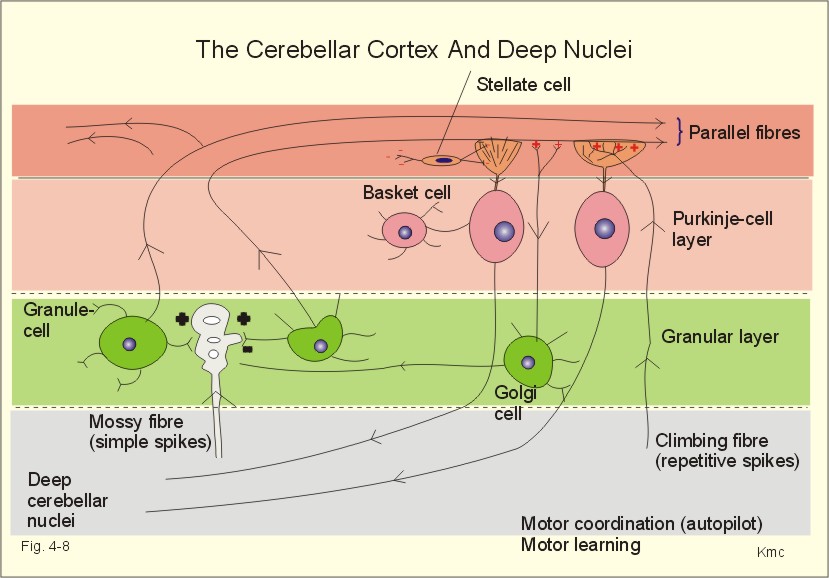
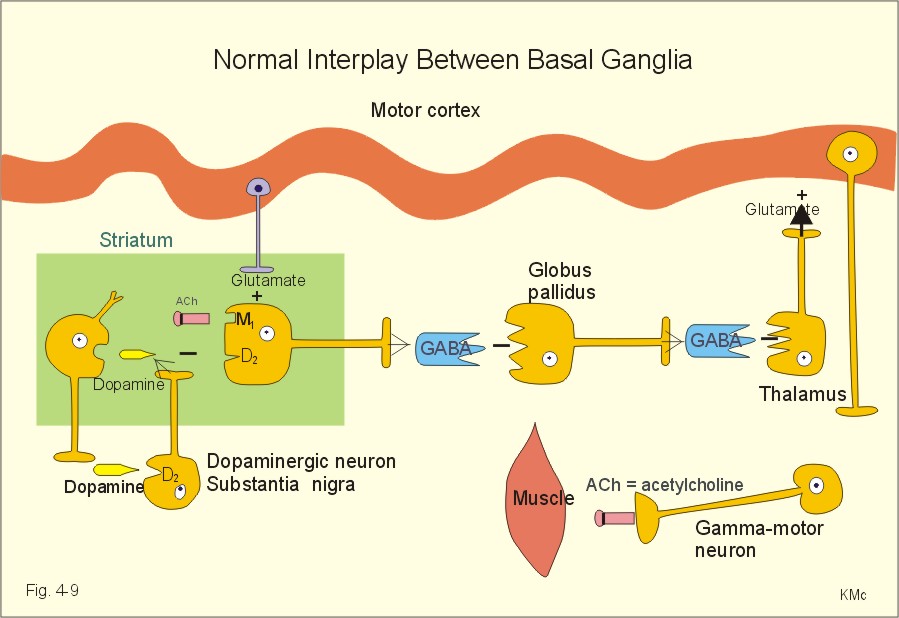
Fig. 4-8: Neuronal connections between the cerebellar cortex and the deep cerebellar nuclei. The cerebellar cortex is build up of three layers. The superficial molecular layer with axons, dendrites and many synapses, the Purkinje-cell layer and the granular layer (Fig. 4-8). The small granule cells send their axons into the molecular layer, where they divide and send so-called parallel fibres in each direction along the folium. These fibres excite the dendrites of the Purkinje and the Golgi cells. The Golgi cells inhibit the granule cells by feedback inhibition. Stellate and basket cells are interneurons that inhibit dendrites and cell bodies of the Purkinje-cells, respectively. Each Purkinje cell is stimulated from a climbing fibre, which projects from the inferior olive. All neurons with cell bodies in the cerebellar cortex are inhibitory except for the granule cells. The cerebellar cortex modulates the activity of the deep cerebellar nuclei. The incoming pathways to the cerebellum end as mossy fibres on the granule cells. Each mossy fibre reach many granule cells. The input signals through the mossy fibres evoke simple spikes (single action potentials) in Purkinje-cells. The climbing fibres produce repetitive or complex discharges in Purkinje cells. Complex spikes of long duration and low frequency are involved in the cerebellar programming of motor learning. The Purkinje-cell axons terminate in the deep cerebellar nuclei or in the lateral vestibular nucleus. This is the basis for cerebellar coordination and fine, rapid adjustments of complex movements. The cerebellar hemisphere affects movements on the same side of the body, because of its crossed connection to the motor system. The motor system projects contralaterally. Discrete electrical stimulation of cerebellum does not cause movements or sense impressions, so it is also termed the silent brain. The vestibulocerebellum projects to the vestibulospinal and reticulospinal tracts, which coordinate balance and eye movements. The vestibulo-ocular reflex produces conjugate eye movements in the direction opposite to that of the head movement. The vestibulo-collic reflex increases the neck muscle tone damping the induced movement. The spinocerebellum receives proprioceptive input from the spinal cord (the spinocerebellar tracts). The spinocerebellum controls the axial muscles through the medial descending motor system, and the proximal limb muscles through the rubrospinal tract of the dorsolateral system. The pontocerebellum receives decision signals and motor control signals from the cerebral cortex by way of pontine nuclei. The pontocerebellum is involved in motor planning, and controls the distal limb muscles through the lateral corticospinal tract. The main function of the basal ganglia is to initiate and stop movements. The basal ganglia inhibit the thalamus, and thus reduce the thalamic stimulation of the motor Cortex.
Fig. 4-9: The basal ganglia and their interplay. Transmitter stimulation is marked by +, and inhibition by -. The affected cell bodies or axons at disease states are marked with a bar. The basal ganglia also contribute to cognitive (i.e., intelligence, knowledge, and motor learning) and affective (i.e., emotional) functions. The basal ganglia include the globus pallidus and striatum. Striatum consists of the nucleus caudatus and the putamen. These deep brain nuclei function in collaboration with several thalamic nuclei, substantia nigra and the subthalamic nucleus (Fig. 4-9). The striatum receives afferent fibres from the cortex (Glutamate + = glutaminergic excitatory fibres), and dopaminergic (inhibitory) fibres from substantia nigra (Dopamine -). Striatum projects to the globus pallidus and to the substantia nigra. These connections are GABAergic and inhibitory (GABA - in Fig. 4-9). Globus pallidus receives afferent GABAergic fibres from striatum, and projects to the thalamus with GABAergic efferents. In the striatum, there are excitatory cholinergic pathways. This paragraph deals with 1.Pure lesion of the medullary pyramid, 2.Abnormal muscle tone, 3. Spinal transection syndrome, and 4. Cerebellar disease. - Capsular stroke, Parkinson´s disease, dyskinesias and epilepsy are all dealt with in Chapter 7, which is a systematic description of neurological and psychiatric disorders. Read chapter 7 before trying to solve the case histories. 1. Pure lesion of the medullary pyramid The control of fractionated finger movements is absent. There is a positive sign of Babinski. Flexion reflexes are not found, and neither is spasticity. On the contrary, muscle tone is decreased. In summary, a pure interruption of the corticospinal tract alone does not show the same signs as capsular stroke. The main deficits caused by medial lesions are reduced muscle tone in the physiologic extensors, loss of balance during walking and standing, and loss of rightening reflexes (they tend to restore head and body position). However, fine finger movements are quite normal. Spasticity is used in clinical neurology to describe muscles resisting fast, passive movements of the limbs, especially in extreme articular positions. When the limbs are moved in extreme articular positions, the increased muscle resistance suddenly disappears. Spasticity includes hyperactive stress reflexes and foot clonus. The resistance dominates in the physiological extensors (antigravity muscles). Spasticity is typical for stroke, where the capsula interna is damaged. The resulting disruption of the lateral descending system is extended by damage of other cortical efferents to the basal ganglia, the thalamus and pons (see Chapter 7). Rigidity is muscle stiffness caused by prolonged activity in the motor units. The muscle resistance is increased towards passive movements of the limbs in any direction (lead pipe rigidity). This condition is found in Parkinson's disease (see Chapter 7). 3. Spinal transection syndrome The spinal shock is immediately recognized by several characteristic symptoms: flaccid paralysis with loss of stretch reflexes, areflexia, loss of autonomic functions, and of all sensation below the level of transection. After a few weeks the spinal shock fades away and the reflexes return and become hyperactive (foot clonus), including mass reflexes and flexion reflexes. A spastic paralysis or paresis replaces the flaccid paralysis. Cerebellum can suffer from damages at two locations: 1. Damage to the flocculonodular lobe causes nystagmus and difficulties in gait and balance (i.e., resembling lesion of the vestibular apparatus). 2. Damage to the vermis or the intermediate region and hemisphere, results in motor disturbances of the trunk and limbs, respectively. Cerebellar disorders include cerebellar incoordination, dysequilibrium, and loss of muscle tone. Cerebellar incoordination comprises ataxic gait, as seen in alcohol intoxication and in disseminated sclerosis. Another type of ataxia is dysmetria, where there is an inability to move the limbs to the desired position. Many patients manifest their ataxia as dysdiadochokinesis, which is a disturbance of the normal ability to make repeated supinations and pronations of the lower arms. Complicated muscle function is stepwise - not smooth. Intention tremor is seen when the patient is asked to touch a target. Speech is slow and slurred, a defect termed dysarthria or scanning speech. Dysequilibrium results in balance problems, and the patient falls to the affected side. Gyratoric vertigo is a genuine rotational or merry-go-round vertigo with the associated loss of equilibrium. This cerebellar vertigo is similar to that following lesion of the vestibular apparatus. Loss of muscle tone is called hypotonia. The hypotonic lack of damping causes the leg to swing back and forth, when the patellar reflex is triggered - so-called pendular knee jerk. Cerebellar nystagmus is involuntary movements of the eyeballs around their natural position - often accompanied by rotational vertigo, when the flocculonodular lobe is damaged. I. Each of the following five statements have True/False options: A. The frontal cortex is involved in motor and emotional behavior. B. The somatic association or interpretation areas (area 5 and 7) are located in the temporal cortex. C. Recycling of information through the primary memory is termed rehearsal. D. Retrograde amnesia following brain commotion is a loss of the short-term memory. E. The limbic system relates behavior and emotions to the intellectual cortex functions. II. Each of the following five statements have False/True options: A. The EEG arousal reaction is a low frequency-high voltage shift. B. Circadian periodicities are changes in biological variables occurring once a day. C. N-methyl-D-aspartate-(NMDA)-receptors bind aspartate, dopamine and glutamate. D. Dreams occur during REM sleep, and the person always reproduces them when awake. E. Dominating EEG frequencies of 8-25 Hz are characteristic of light sleep. III. Each of the following five statements have True/False options: A. Fast fatigable motor units consist of type IIB twitch fibres with few mitochondria and small amounts of myoglobin. B. The Renshaw cells synapse with a-motor neurons of antagonistic muscles, and thus inhibit monosynaptic reflexes. C. The cerebellar hemisphere affects movements on the opposite side of the body. D. The Golgi tendon organs are the serially located terminals of group Ib fibres wrapped around bundles of collagen fibres in the tendons. E. The ventromedial descending system involves the ventral corticospinal tract and much of the corticobulbar tract ending in the medial group of brainstem and spinal cord interneurons. An outstanding Russian composer, 63 years of age, recovered from a cerebral insult. However, he could no longer understand spoken or written language, although his speech was fluent. The composer also maintained his ability to compose excellent music. 1. What is the name of this deficit in language function? 2. Where in the brain is the lesion localized and in what side of the brain? A male of 65 years suddenly falls and is found in deep coma by the doctor. There is a left-sided hemiplegia with short arm-long leg as a flexion reflex. The paralysis and areflexia turns into spastic hemiparesis with a positive sign of Babinski. The deep stretch reflexes (patellar- and Achilles-tendon reflexes) are enhanced. There is loss of superficial reflexes (the abdominal and cremasteric reflexes). When the Achilles-tendon reflex is triggered it releases foot clonus. When the patient is awake from coma his facial nerve paresis is examined. He can knit his brows and turn his eyes upwards. 1. What is the pathophysiologic basis for this condition? 2. What are spasticity and foot clonus?
Try to solve the problems before looking up the answers .
Schuman, E.M., and D.V. Madison. "Nitric oxide and synaptic function." Annu. Rev. Neurosci. 17: 153-183, 1994. Thomson, R.F. "The brain. A Neuroscience Primer." Third Edition. Freeman, 2000. McIntosh, A.R., C.L. Grady, L.G. Ungerleider, J.V. Haxby, Rapoport, S.I., and B. Horwitz. "Network analysis of cortical visual pathways mapped with position emission tomography." J. Neurosci. 14 (2): 655-666, 1994.
|
||









.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Click here to introduce your comments or contributions