New Human Physiology | Paulev-Zubieta 2nd Edition
Chapter 29: Sexual Satisfaction, Reproduction and Disorders
| HOME | PREFACE | TABLE OF CONTENTS | SYMBOLS | SECTION INFO | CONTRIBUTORS | LINKS | CONTACT US |
Highlights
Study_ObjectivesPrinciplesDefinitionsEssentials
PathophysiologyEquationsSelf-AssessmentAnswers
Further Reading
|
Chapter 29
|
 |
|
|
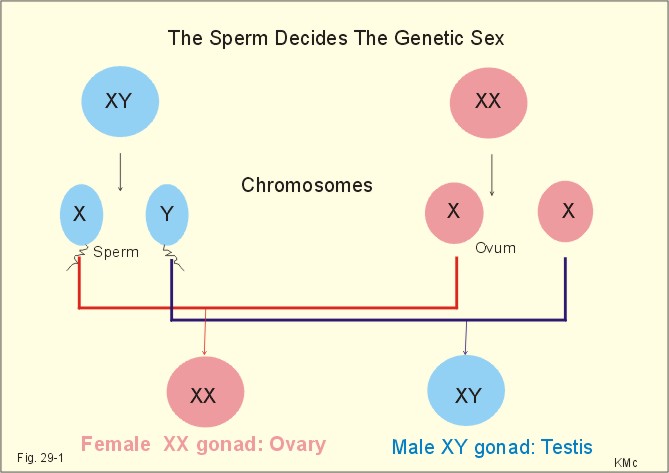
· To define amenorrhoea-oligomenorrhoea, dyspareunia, gynaecomastia, hypogonadism, impotence, infertility, menarche, menopause, menstruation and phases of the menstrual cycle, oligospermia-azoospermia, sterility, and virilization. · To describe anticonception, anovulatory cycles, bleeding disturbances, castration, cryptorchism, postmenopausal hormonal alterations, puberty, anabolic steroids and doping, genetic and psychosocial sexual disorders. · To explain the effect of anabolic steroids, the normal menstrual cycle, conception, implantation, pregnancy, pregnancy tests, birth and suckling. To explain the normal ovarian and testicular function, gametogenesis, erection, ejaculation and sexual satisfaction (orgasm). To explain the effect of androgen-binding protein, inhibin, aromatase, and the biosynthesis of steroids. · To use the above concepts in problem solving and case histories. · The gonads are concerned with the well being and preservation of the human race. · The sperm decides the genetic sex (genotype). The Y chromosome is a constant determinant of maleness. · Foetal differentiation of the genital ducts and of the external genitalia requires foetal gonadal hormones. The foetal genital tract will always develop into female genitals, if unexposed to embryonic testicular secretion. · Amenorrhoea-oligomenorrhoea are terms used for absence - irregular, infrequent menstrual periods. These signs suggest female hypogonadism, when pregnancy is excluded. · Azoospermia describes absence lack of sperm in the ejaculate · Dyspareunia refers to female pain or discomfort during intercourse. · Gametogenesis is the formation of ova and sperm. The primitive germ cells are divided by meiosis, so the number of chromosomes is halved (22 autosomes and one sex chromosome). · Genetic sex is determined by the presence or absence of the Y chromosome. The Y chromosome determines the development of testes and maleness. The Y chromosome contains a sex determining region (the SRY gene), which encode the testis determining factor (TDF). · Genital sex is the phenotypic sex (apparent female or apparent male). · Gonadal sex is determined by the presence of normal ovaries or testes. · Gynaecomastia refers to the occurrence of female breasts in males. The causes are HCG-producing tumours, oestrogens or oestrogenic drugs. · Hypogonadism (male) refers to a condition with small, soft testes producing little sperm and testosterone. The condition is usually found with subfertility. · Impotence is inability of the male to produce an adequate erection for satisfactory sexual intercourse. · Infertility (subfertility) is a diagnosis used on a couple, which has been unable to conceive during one year of unprotected intercourse. The causes are oligospermia, tubule blockage, ovulatory disorders, or combined problems with both persons in the couple. · Menarche refers to the age at the first menstrual period. · Menopause refers to cessation of periods, which usually occurs around the age of 50 years. · Menstruation is the onset of spontaneous regular uterine bleeding. · Oligospermia refers to reduced numbers of sperm in the ejaculate. The causes are primary testicular disease or blockage of the vas deferens. · Puberty is the transition period from a non-reproductive to a reproductive state. · Sterility refers to individual infertility. Chemotherapy and other drugs may cause sterility. Surgical blockage of the tuba or the vas deferens results in sterility. · Virilization is the occurrence of male secondary sex characteristics in the female. · Definitions of other genetic concepts are given in Chapter 31. This paragraph deals with 1. The sexual drive , 2. Sex before birth, 3. The menstrual cycle, 4. Ovulation/Female orgasm, 5. Conception, 6. Breast development, 7. Labour, 8. Efferent activity during coitus, 9. Sex hormones, and 10. Male puberty. We feel the sexual drive or desire for sex (libido), when sex-related areas in the higher brain centres are stimulated. These centres include the limbic system, Stria terminalis and the preoptic region of the hypothalamus. The desire for sex is increased by androgens in both sexes. The sex desire of females is variable - for some it increases near the time of ovulation, when oestradiol secretion is increasing, while others experience a peak drive near menstruation. The CNS cells involved (see above) must contain sex hormone receptors. Sex hormones are steroids. They are lipid soluble and pass the cell membrane easily. After binding to cytoplasmic receptors (the steroid-thyroid family), the receptor-hormone complex translocates to the cell nucleus. Here the information is transcribed and translated. The result is release of new proteins with the same information into the cytosol, where the physiologic response is triggered. Castration is assumed to reduce female libido minimally, but male libido is most often lost. Removal of one testis need not change the male libido. These clinical observations reflect psychosocial differences, and not necessarily a different libido mechanism in the two sexes. Hypothyroid persons lose their sex drive. The sex desire (libido) is stimulated by a multitude of sense impressions (visual, auditive, olfactory, and psychological). Potency refers to the ability to engage in intercourse. The brain is an important sex organ. Obviously, any natural body contact can be considered part of a healthy sex life - including the penetration of the penis in the vagina. Sexual satisfaction is synonymous with orgasm in Western cultures. Orgasm is the psychological climax or the culmination of total commitment in a sexual act that is accompanied by a series of physiologic reactions. Female orgasm involves spinal cord reflexes similar to those involved in male ejaculation (see later). One very important reaction is ovulation, which is an automatic consequence of copulation among many animal species and periodically in humans. Sexual enjoyment covers several phenomena. For example the fetishist satisfaction of wearing the clothes of the opposite sex. This is the important part of a transvestites sex life. Some transvestites become asexual in the general sense of the term, since they do not need partners. Some individuals prefer masturbation (onany) as a substitute for partnership. Many individuals prefer heterosexual contacts; others prefer homosexual activities, while bisexuals may prefer either sex - depending on the circumstances. Sexual activities can vary. Besides, homosexual activity, oral sex, anal sex and many other variants are not uncommon. Normal sexual development in the embryo involves several processes. The sperm, which can be an X or an Y chromosome sperm, decides the genetic sex or sex genotype (Fig. 29-1). The genetic sex is independent of the ovum. If the ovum is fertilised by an X spermatozoa (22 + X-chromosomes) the offspring is XX, a female. If the ovum fertilises by an Y spermatozoa (22 + Y-chromosomes) the offspring is XY, a male (Fig. 29-1).
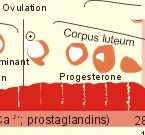
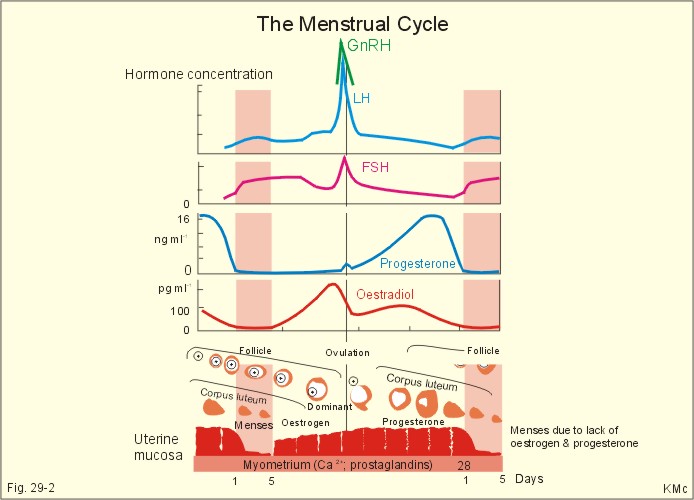
Fig. 29-1: The sperm decides the genetic sex. The presence of the Y chromosome is the determinant of maleness. Sex differentiation in the embryo usually harmonises with the sex genotype, but hormonal disturbances can lead to abnormalities. Proliferation of non-germinal and germinal cells in the genital ridge creates the gonadal primordia, which develops into a cortex surrounding the medulla. Until the 7th week of gestation, each sex has a bipotential system (the sexual indifferent stage) with both Wolffian and Müllerian ducts. The urogenital sinus develops into the external genitals in both females and males. Around the 7th week, the medulla of the primitive gonad begins to differentiate into a testis, if an Y chromosome is present. This is because the Y chromosome contains the so-called SRY gene (the sex determining region of Y), which encodes the testis-determining factor. As the testes grow and their Leydig cells start to produce testosterone, the Wolffian ducts develop into the male reproductive tract (epididymis, vas deferens, seminal vesicles and the ejaculatory ducts), whereas the Müllerian ducts regress. Testosterone stimulates the growth and differentiation of the Wolffian ducts in the male. The regression of the Müllerian ducts is caused by the antimüllerian hormone from the Sertoli cells. Conversely, in the female, the cortex of the indifferent gonads differentiate into ovaries, if only two X chromosomes are present and no Y. In the female foetus, where there is a developing ovary and no antimüllerian hormone, the Müllerian ducts develop into the female reproductive tract (the uterine tubes, uterus and the upper vagina), and the Wolffian ducts degenerate because the ovary does not secrete testosterone.- When a normal female foetus is exposed to androgens during the period of differentiation of the external genitalia, an apparent male can result. Visible differentiation of the gross anatomy does not appear until late in the second month of embryonic life. Testosterone causes the differentiation of the foetus to a male. The foetal genital tract will always develop into female genitals, if unexposed to embryonic testicular secretion. The genital sex is a phenotypic female. If testosterone is present, male external sex organs develop and the genital tubercle elongates to form the male phallos. If testosterone is absent, female organs develop instead. It is the action of testosterone and 5-a-dihydrotestosterone on the urogenital sinus that is behind the normal development of the male external genitalia. In the last months of gestation the growth of the external genitalia depends upon foetal pituitary LH. One population of cells in the indifferent gonade develops into the granulosa cells of the ovarian follicle and the Sertoli cells of the testicular seminiferous tubules. These cells support and mature the germ cells. – Another population of so-called interstitial cells develop into the theca cells of the ovary and the Leydig interstitial cells in the testis. The Leydig interstitial cells secrete testosterone, in response to human chorionic gonadotropin (hCG) from the placenta. The presence of normal ovaries or testes determines the gonadal sex. Without normal ovaries or testes any genetic sex will develop into an apparent female. Foetal plasma growth hormone (GH) concentrations are high, but GH-receptors are deficient and foetal GH is not essential for linear growth. Prolactin and placental GH act as growth factors and induce the presence of IGF-1 and IGF-2. A small transfer of maternal thyroid hormone is important for early foetal development. At birth, the babys own thyroid hormone is important for CNS development and somatic growth. Foetal PTH stimulates the Ca2+-transfer across the placenta and controls plasma-Ca2+. Foetal ACTH is important late during gestation in particular at birth, and the cortisol concentration is high in umbilical cord plasma. Foetal pancreatic a-and b-cells are functional by 14 weeks of gestation, but their release of glucagon and insulin is low. In 1949 Barr et al. found a densely coloured body in the periphery of the nucleus (the Barr body or sex chromatin) of the buccal mucosa of females. The Barr body is also present in other individuals with two or more X-chromosomes in each cell. Individuals with one sex chromatin (Barr body) also have a drumstick attached to a small fraction of their leukocytes (Fig. 29-6). We find sex chromatin and drum sticks in cells, whether they divide or not. Chromosomes are only visible in dividing cells. The maximum number of sex chromatin and drumsticks is always one less than the number of X-chromosomes (Fig. 29-6). The menarche is the age at the first menstrual bleeding. It often occurs between the 12th and the 14th year. LH and FSH are coordinators of gonadal function. The secretion of these pituitary gonadotropins is regulated through negative feedback by the plasma concentration of gonadal steroids. LH stimulates the interstitial cells of the ovaries (and testes), but LH also acts on female granulosa cells. LH binds to a LH-receptor, which spans the cell membrane several times. The LH receptor acts via adenylcyclase and with cAMP as a second messenger. Prostaglandins may increase the cAMP effects. Maintained stimulation by LH down-regulates the number of LH-receptors on the surface of gonadal cells. FSH acts on ovarian granulosa cells (and testicular Sertoli cells) by binding to FSH-receptors, partially homologue with the LH-receptors. The increase in cAMP following FSH-receptor binding transcribes the aromatase gene and stimulates oestrogen synthesis. FSH stimulates synthesis of inhibin and peptide/protein products from granulosa and Sertoli cells. FSH amplifies the sensitivity to LH by increasing the number of LH-receptors on granulosa cells. LH and FSH increase glucose oxidation, lactic acid production and protein synthesis. The menstrual cycle starts at the first day of bleeding (menstruation). The bleeding is due to decrease of oestrogen and progesterone secretion. The FSH and LH secretion start to rise and stimulate the growth of several follicles ‑ in particular following the bleeding. One of these – the dominant follicle – select itself by outstripping the others and grow so fast that the follicle can protrude more than 10 mm from the surface of the ovary. The dominant follicle has an increased oestrogen synthesis due to increased aromatase activity. Oestrogen from the granulosa cells of the dominant follicle binds to specific, cytoplasmic receptors (of the steroid-thyroid-family) in the endometrial and other uterine cells. Oestradiol activates and stimulates formation of oestrogen and progesterone receptors.
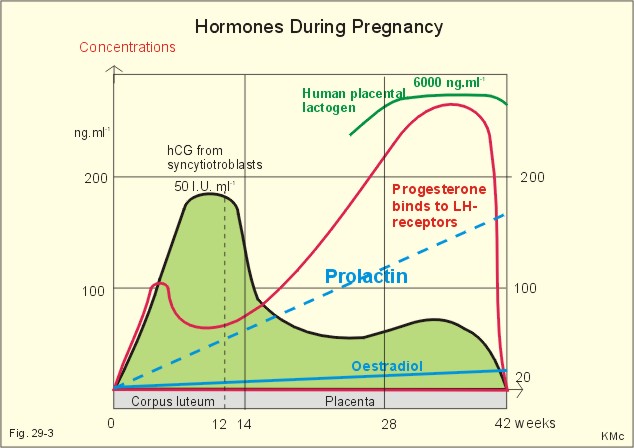
Fig. 29-2: The menstrual cycle in a female. Oestrogen increases the thickness of the endometrium, the size of the myometrial cells and the number of gap junctions thus allowing the myometrium to work as a unit. The oestrogen phase is also called the proliferative phase. The concentration of sex hormones in plasma is shown in Fig. 29-2. Oestrogens work synergistically with progesterone to release gonadotropins by positive feedback just before ovulation. Following the rupture of the follicle (ovulation), the corpus luteum produces increasing amounts of progesterone in addition to oestradiol also from a new developing follicle (Fig. 29-2). Due to the priming effect of oestrogen on progesterone receptors, both hormones stimulate the growth of the endometrial glands, so that they curl like a helix. The progesterone effect in particular provides the endometrial/myometrial tissues with their high secretion and bloodflow, so the uterus is prepared to receive the fertilised ovum. During sexual stimulation the vaginal fluid secretion increases, as does the bloodflow of the organs involved. If fertilisation does not occur, the level of oestradiol and progesterone switches off both gonadotropins. The corpus luteum fades out and degenerates with no LH to support it (Fig. 29-2). The ovarian hormones almost cease to flow, and the uterus is deprived of their stimulating action. Therefore the uterus shrinks and sheds its swollen lining. On the first day of the menstrual bleeding, the low progesterone and high prostaglandin level probably releases enough Ca2+ to start spontaneous contractions of the myometrial cells. Ca2+ -ions enter myometrial cells and stimulate their activity in the secretory (progesterone) phase. The gap junctions synchronise these contractions, so that they include the whole myometrium. This can make excretion of blood and necrotic cells (containing prostaglandins) extremely painful. Prostaglandins dominate in menstrual fluid and stimulate the spontaneous activity of the human myometrial cells. A normal bleeding corresponds to a loss of up to 50 ml of whole blood. The mixture of vaginal fluid and menstrual blood produces a pH close to that of normal blood. The average cycle length is 28 days. ADH (vasopressin) secretion from the neurohypophysis can cause pre-menstrual tension and an unpleasant increase in body fluid volume. OvulationA sudden increase in the plasma level of oestradiol maintained for more than 24 hours can increase FSH output by positive feedback. This is called the positive feedback release ovulation. The pulsatile release of GnRH from the hypothalamus is possibly stimulated by the high oestradiol concentrations in mid-cycle and oestradiol increase the number of GnRH receptors on the gonadotropic cells of the anterior pituitary. A neural hypothalamic pulse generator has been proposed to be involved in ovulation, and in some cases female orgasm triggers ovulation. At lower plasma levels oestradiol is a potent inhibitor of GnRH secretion and thus of FSH and LH output (negative feedback). The negative feedback forms the basis for the ovulation-inhibition by contraceptives. LH binds to a membrane LH-receptor and acts via a G-protein, adenylyl cyclase and cAMP. LH mobilises cholesterol and its conversion to progesterone. FSH acts on ovarian granulosa cells and testicular Sertoli cells by binding to a membrane receptor homologous with the LH-receptor. The binding increases the transcription of the aromatase gene, the oestrogen and the inhibin synthesis. The primary inhibitor of FSH secretion is the peptide, inhibin, that is secreted by the ovary (and testis), and blocks the effect of GnRH. The oestradiol release from the dominant follicle increases sharply in the last part of the follicular phase. This triggers the preovulatory surge of gonadotropins (LH and FSH). The LH surge induces an enzyme that increases the synthesis of leukotrienes, prostaglandins and thromboxanes. These molecules create an inflammation that causes rupture of the follicle. LH continues to act on the follicular granulosa cells, turning them into a yellow endocrine organ, the corpus luteum. OrgasmThe time for preplay including clitoral and multifocal stimulation is important for most females. A clitoral orgasm in the preplay often triggers more female orgasms later during the intercourse. Female orgasm is released from the spinal cord reflexes via sympathetic signals in the pudendal nerves. Two persons with a simultaneous sexual drive must have the necessary time for the sexual act. If they are also in love, it is natural to explore and use all means to satisfy each other. Years ago, when the Kinsey report was made, the average duration of sexual intercourse was measured in seconds in the US. American males able to ejaculate even faster were assumed to be particularly virile. Today, such a short performance is considered a male disease called premature ejaculation. Conception Approximately 100-200 million sperms are produced each day of the fertile lifespan. The female foetus may contain 6 million oocytes, but the number decreases throughout her life (less than half a million at puberty and she may have 500 ovulations before the menopause). The autonomic moving spermatozoa passes through the uterus while prostaglandins inhibit their spontaneous activity. The spermatozoa can keep their vitality for more than 2 days, if they reach the fallopian tube. They lose their protective cover in the fallopian tube. The head of the spermatozoa swell and liberates proteolytic enzymes. These enzymes can dissolve the zona pellucida around the egg (oocyte). All these events in the spermatozoa takes days before it meets with the oocyte. The oocyte can only live 12-24 hours without conception. Pregnancy Many sperms bind to the zona pellucida, but only one penetrates the wall – and blocks the entry of other sperms. Fusion of the two sex cell membranes forms the zygote, and the mitosis is complete within 24 hours. The zygote passes into the uterine tube within a few days, protected against other spermatozoa by an increased permeability for K+, so that the zygote membrane hyperpolarises. Peristaltic movements of the tube and ciliary motion conduct the zygote to the uterine cavity while undergoing cleavage division. Each cell is capable of developing into a complete human being up to the eight-cell stage. At the morula stage, the cells start to develop into the inner cell mass or blastocyst, and the trophoectoderm or trophoblast. Seven days after conception, the blastocyst loses the zona pellucida and implants in the wall of the uterus (nidation). Nidation depends on prior conditioning of the endometrial stromal cells by progesterone bringing it into the proliferative phase. The stromal cells accumulate nutrients and swell or decidualize around the blastocyst. Endometrial laminin and fibronectin facilitate adhesion. Histamine and prostaglandins increase the permeability of the vessels around the nidation site. More than 2/3 of all conceptions result in miscarriage, because of insufficient attachment or other anomalies. The foetal trophoblast, which give rise to the extra-embryonic tissues differentiates into two cell types. An inner layer of cytotrophoblasts, and an outer layer of syncytiotrophoblasts. The cytotrophoblasts synthesise stimulatory hormones such as CRH, GnRH, TRH and steroids. The syncytiotrophoblasts synthesise first of all human chorionic gonadotropin (hCG). The b-group of hCG is specific and detected in maternal plasma 6 days following conception by specific antibody methods. The hCG is detectable in the urine within 9 days after conception. The placenta is a fantastic hormone factory, which produces large amounts of hCG, relaxin. oestradiol, progesterone and human chorionic somatomammotropin (hCS or human placental lactogen, hPL). The hPL is synthesised from the 4. week of gestation. The hPL stimulates maternal lipolysis and inhibits insulin effects, causing hyperglycaemia. The hCG is chemically related to TSH, FSH and LH. The hCG acts like LH and binds to the LH-receptors. The secretion of hCG is stimulated by GnRH produced by cytotrophoblasts. This is what keeps corpus luteum in being, and the pregnancy continues. During pregnancy, hCG thus conserves the corpus luteum, taking over the role of LH. The secretion of hCG stimulates ovarian release of progesterone and oestrogens just like LH. The hCG stimulates production of relaxin, inhibits the maternal secretion of LH and stimulates the maternal thyroid gland causing struma or hyperthyroidism in some pregnant females. Inhibin A from foetal trophoblasts peaks within the first week and suppresses maternal FSH secretion. Inhibin B concentrations remain low throughout gestation. LH and FSH concentrations in foetal plasma peak in mid gestation.
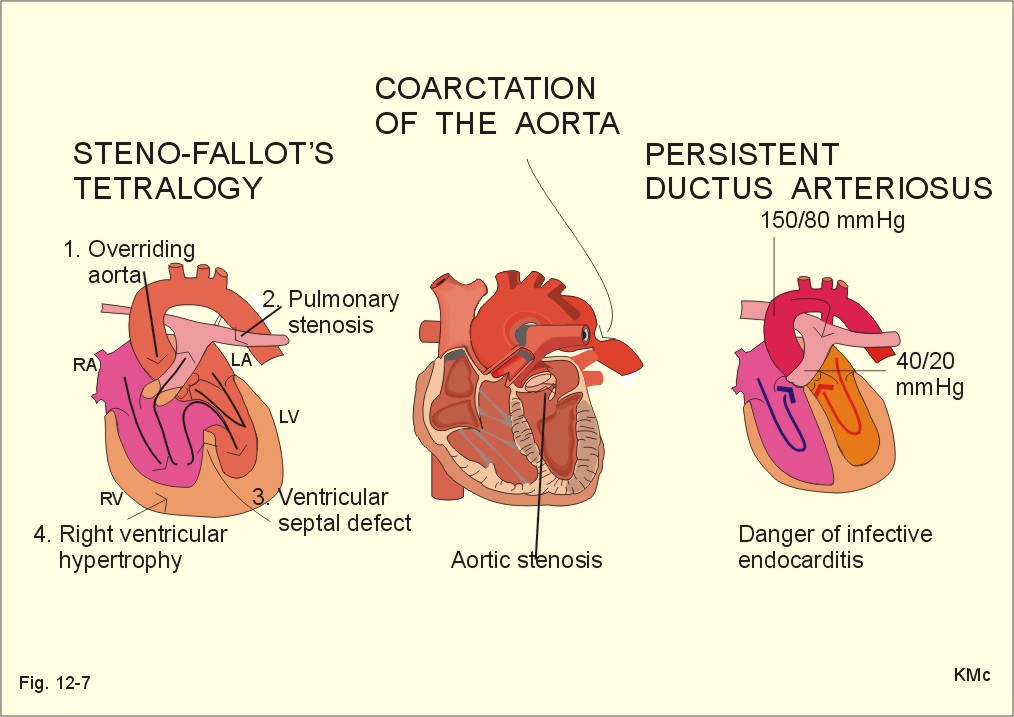
Fig. 29-3: Variations in plasma hormone concentrations during a normal pregnancy (42 weeks). The plasma [hCG] reaches a peak value after 10 weeks of pregnancy, when the syncytiotrophoblast count is maximum (Fig. 29-3). - Shortly after delivery hCG disappears. The first peak on the plasma progesterone curve is progesterone produced by corpus luteum. The placenta takes over the progesterone production during the remaining pregnancy period ending with a peak concentration before birth. Progesterone protects the foetus in the uterine cavity by stimulation of endometrial glands that nourish the zygote and by maintenance of the decidual cells. Progesterone inhibits uterine contractions (inhibits prostaglandin synthesis and oxytocin sensitivity). The foetus and the placenta form a foetoplacental unit. It produces all the hormones necessary for a successful pregnancy. Steroid precursors are delivered from both the foetus and the mother. Oestrogen (oestradiol, oestrone, and oestriol) concentrations rise steadily throughout pregnancy (Fig. 29-3). Oestrogens stimulate the growth of the myometrium and of the ductal system of the breast. Oestriol production independs on the foetal adrenal cortex, so maternal plasma oestriol provides an estimate of the foetal condition. The placental progesterone blocks the menstrual cycle of the mother. Pregnant females therefore develop amenorrhoea. During puberty FSH, LH, growth hormone, and insulin are important for the breast development. The thyroid hormones (T3/T4) are permissive. Before puberty, plasma LH and FSH concentrations are low. There is no reaction to the low concentrations of gonadal steroids and inhibin. Oestrogens are growth factors for the myometrium and for the ductal system og the breast during pregnancy. At the end of pregnancy there are other hormonal events. Progesterone secretion reaches a peak and then falls. This fall in progesterone allows the pituitary to release prolactin (LTH). Prolactin from the maternal pituitary rises throughout pregnancy. Prolactin acts on the enlarged mammary glands turning them into milk producers. Prolactin develops the milk producing acini in the breasts during pregnancy. Prolactin Inhibiting Factor (PIF or dopamine) from the brain inhibits the prolactin secretion. The baby’s suckling stimulate the secretion of prolactin and oxytocin, but oestradiol and sexual stimulation is also involved. The mechanical stimulation of the breast increases the secretion of prolactin from the pituitary, but the response is strikingly reduced by alcohol. Prolactin is important for the development of the mammary gland tissue, oxytocin, however, governs the ejection of milk during lactation. Oxytocin causes contraction of the myoepithelial cells in the milk ducts (just as it does in the myometrial cells). Mother-milk contains long chain fatty acids that are essential for brain development. Suckling babies are protected against juvenile diabetes in comparison to non-suckling babies. Cow's milk contains much more protein and less lactose than human milk. When the foetus has reached a critical size, the myometrial fibres are stretched, which increase their contractility. At the end of pregnancy the uterus is sensitised by oestrogen. After a high peak in progesterone secretion the progesterone output falls. This fall in progesterone allows the uterus to respond to oxytocin, whose release is the final trigger for parturition. The foetal pituitary-adrenal axis signals to the placenta a decrease in the progesterone-oestrogen ratio acting on the myometrium. This increases myometrial contractions that are mediated by prostaglandins (PGE2 and PGE2-a). A local increase in prostaglandin concentration increases myometrial cell Ca2+ and triggers uterine contractions. The density of oxytocin receptors in the myometrium increases throughout gestation and particular at term. The role of the stable plasma concentration of maternal oxytocin at parturition is an enigma. Oxytocin is released according to a pulsatile pattern. The frequency of oxytocin pulsations increases at labour. This fact does not exclude an important role of oxytocin in normal human parturition. Therapeutic doses of oxytocin initiate labour in most cases at the end of gestation, The foetal cortisol production prepares the foetus to adapt to extrauterine life by stimulating lung maturation, by increasing the hepatic glycogen stores, and by promoting closure of the ductus arteriosus (Fig. 12-7). Relaxin is an insulin-like polypeptide produced by the corpus luteum and placenta. The hormone relaxes pelvic articulations, suppresses myometrial contractions and softens the uterine cervix in order to facilitate passage of the foetus. Several other factors are involved in human labour, but the exact trigger mechanism remains unclear. 8. Efferent activity during coitus The activity in males is described as an example. The typical sequence of efferent events in the male includes erection, emission of semen and ejaculation. Erection means penile rigidity and elongation due to parasympathetic vasodilatation. Psychological factors trigger penile rigidity, and sexual thoughts can cause erection, emission and ejaculation. The penis contains erectile tissue located in two dorsal corpora cavernosa and in a single ventral corpus spongiosum. All the cavernous spaces of the three penile corpora receive blood from thick-walled arteries ending centrally in each corpus. The blood leaves the cavernous spaces through thin-walled veins starting peripherally. Tactile stimuli, especially from the very sensitive glans penis activate sensory, somatic fibres in the pudendal nerve, whereby impulses reach the sacral plexus. Parasympathetic impulses (S2-S4) from the sacral plexus elicit dilatation of the arteries and constriction of the veins in penis. The cavernous spaces are hereby filled with blood under high (arterial) pressure within seconds, causing the penis to become hard and elongated for penetration. - Erection occurs quite normally during the REM phases of sleep. Emission is caused by sympathetic contraction of smooth muscles (in epididymis, ducts and glands), which drive the fluids into the posterior urethra. Oxytocin ejects sperm into semen. Two exocrine glands near the neck of the bladder (the seminal vesicles and the prostate) secrete fluids that nourish the sperm and transport it through the urethra during the sexual act. The prostate gland supply an alkaline secretion containing Ca2+, Zn, and phosphatase to the ejaculate. The seminal vesicles supply fructose and prostaglandins. These two secretions neutralise the acid semen and help propel the spermatozoa towards the ovum. Seminal fluid also contains gonadotropins, sex hormones, inhibins, endorphins, relaxin, proteases and plasminogen activator. Epididymis supply sperm-coating proteins. Ejaculation: Ejaculation is a sympathetic response. Contractions of skeletal muscles expel the semen from the urethra in a rhythmic pattern. Signals from glans penis reach the lumbar region of the spinal cord through afferent fibres in the internal pudendal nerves. Filling the posterior urethra with semen triggers sensory impulses that travel through the pudendal nerves to the spinal cord. The spinal cord transmits rhythmic signals to the skeletal ejaculation muscles (the ischio-and bulbo‑cavernous muscles and those of the pelvis). These rhythmic signals stimulate rhythmic contractions that expel the semen from the urethral meatus into the female genitals. – A typical ejaculate contains 300 million spermatozoa in 3 ml. Inside the female genitalia the sperm is subject to the process of capacitation, which takes place within 6 hours. The sperm head is coated with substances from the ejaculate, Ca2+ enters the sperm, sperm motility increases, and the ability to penetrate the ovum is enhanced. The acrosomal membrane fuse with the outer sperm membrane, so that pores are formed and proteolytic enzymes can reach the surface of the sperm head. Sex hormones are oestrogens, progesterone, androgens and eichosanoids. Steroid synthesis in the gonads begins with cholesterol from acetyl Coenzyme A, and is almost identical to that of the adrenal cortex. Oestrogens stimulate the female genitals and act to produce female secondary sex characteristics when a female enters puberty. Oestrogens and progesterone all enters the cell cytosol easily and bind to cytoplasmic receptors of the steroid-thyroid family. Oestradiol increases the synthesis of oestrogen- and progesterone-receptors. These sex characteristics include the progressive growth of fallopian tubes, uterus, vagina, and external genitalia; also the fat deposition in breasts, buttocks, and thighs (Fig. 29-4). The ductal and stromal growth of the breasts is initiated just as the general growth at puberty with increased RNA and protein synthesis in the body cells. Oestrogens stimulate secretion of prolactin from the pituitary lactotrophic cells, increase the thickness of the endometrium and the size of the myometrial cell and their number of gap junctions. Oestrogens stimulate the hepatic production of essential proteins (eg, TBG, blood clotting factors, plasminogen, and HDL), but they inhibit formation of antithrombin III and LDL. Retention of salt and water can cause oedema. Oestrogens consist of oestradiol, the principal ovarian oestrogen, oestriol, the major placental oestrogen, and oestrone, an important ovarian and postmenopausal hormone. At a certain level oestradiol increases GnRH secretion and FSH output by positive feedback. There is also an increased LH sensitivity to GnRH. This feedback is already called positive feedback release ovulation, where the leading follicle ruptures. At lower oestrogen levels in the blood, it is a potent inhibitor of gonadotropin releasing hormone (GnRH) secretion and thus of FSH output. This is the reason for the ovulation-inhibition by many oral contraceptives. In the blood oestradiol is bound to sex steroid-binding globulin.
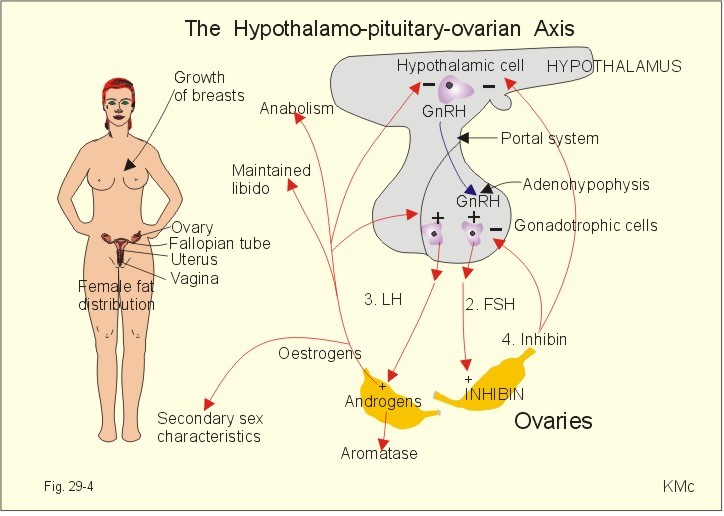
Fig. 29-4: Feedback loops and targets organs in the hypothalamic-pituitary-ovarian axis. The hypothalamic GnRH secretion shows a cyclic variation in adult females of approximately 28 days, probably a genetic code imposed by the CNS. 1. Peaks of GnRH release reach the adenohypophysis through the portal system, and release both FSH and LH to reach the ovary via the systemic circulation (Fig. 29-4). 2. FSH stimulates follicular growth, inhibin-release from stromal cells, and aromatase activity in the ovary. Aromatase converts ovarian and other androgens to oestrogens. 3. LH stimulates the ovarian androgen production. 4. Inhibin is the primary inhibitor of FSH release by blocking the effects of GnRH on the adenohypophysis. Oestrogens are responsible for the female secondary sex characteristics, the maintenance of libido, anabolic effects, and the negative feedback on the GnRH secretion and the Gonadotropin secretion of the adenohypophysis (Fig. 29-4). Progesterone secretion rises sharply in the luteal phase of the menstrual cycle, and modulates the effect on oestrogens on the endometrium and the myometrial cells. Since the oestrogens have primed all the progesterone receptors, both hormones stimulate the growth of endometrial glands so they curl. Progesterone stimulates the secretion and high bloodflow of the uterus, so it is prepared to receive the fertilised ovum. Progesterone increases the basal core temperature by 0.5 oC, which is used as an indicator of ovulation. In the absence of pregnancy, progesterone secretion falls and switches off the release of GnRH and both Gonadotropins (Fig. 29-4). The corpus luteum degenerates, resulting is sloughing of the endometrium (ie, menstruation). Pregnancy is maintained by progesterone, and uterine contractions are inhibited. Progesterone has a certain aldosterone effect by competition for the same receptors. Progesterone has a negative effect on the lipid profile by increasing the LDL and reducing the HDL fractions in the blood plasma. Androgens, such as testosterone, are anabolic, maintains spermatogenesis and libido, and act to produce male secondary sex characteristics. These characteristics are the deepening of the voice at puberty, beard, body hair, sebaceous glands in the skin, as well as the growth of the skeleton, the striated muscle system, the external genitalia and male behaviour-attitude. The primary sex structures are the testes with seminiferous tubules, epididymis, prostate, and seminal vesicles. Testosterone is responsible for the growth, maturation, and maintenance of the primary sex structures. Androgens stimulate the growth and polyamine synthesis in the prostate and the seminal vesicles. Hereby RNA synthesis is stimulated and the result is often hypertrophia and hyperplasia. Testosterone increases LDL and decreases HDL concentrations in plasma.
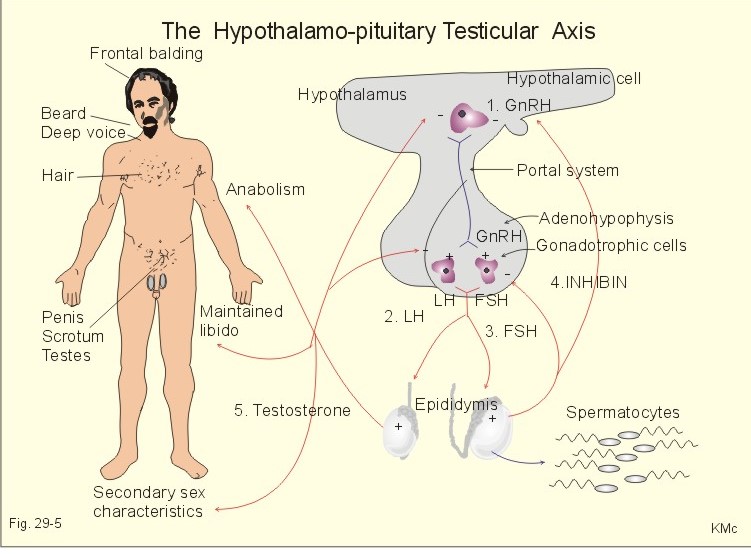
Fig. 29-5: The feedback system of the hypothalamic-pituitary- testicular axis. Testosterone is reduced to two other potent androgens (dihydrotestosterone, 5a-androstendiol) in many tissues. Testosterone is thus a prohormone for these potent androgens. Most of the testosterone in plasma binds to sex steroid-binding globulin, a small fraction binds to albumin and only 1% is free testosterone. Thyroid hormone and oestrogens increase the concentration of sex steroid-binding globulin and thus reduces the free fraction. Androgens have the opposite effect. Testosterone diffuses easily into the cell cytoplasm and binds to a cytoplasmic receptor belonging to the steroid-thyroid receptor superfamily. The male sexual system is controlled in the following way: 1. GnRH (= LHRH) is released from hypothalamic cells in a pulsate pattern, and stimulates release of LH and FSH from gonadotropic cells of the adenohypophysis (Fig. 29-5). 2. LH stimulates the Leydig cells of the testes to produce testosterone. These cell also produce small amounts of oestrogens, oxytocin and subunits of pro-opio-melanocortin. 3. FSH and testosterone stimulate the Sertoli cells of the testicular seminiferous tubules to produce spermatocytes and inhibin. 4. Inhibin is a glycoprotein that reduces the pituitary FSH secretion (blocks the effects of GnRH) by negative feedback (Fig. 29-5). Activins are synthesized by subunits of inhibin. The Sertoli cells produce inhibin, as do the granulosa cells in females. Inhibin inhibits FSH but not LH secretion by the pituitary gland. Activin stimulates FSH secretion just as GnRH. Follistatin binds and neutralises activin, so follistatin inhibits FSH-secretion. 5. Testosterone is responsible for the male secondary sex characteristics, the maintenance of libido, anabolic effects, and the negative feedback on the GnRH secretion and the Gonadotropin secretion of the adenohypophysis. Acne during puberty is due to testosterone, but in the female adrenocortical androgens are involved. Testosterone promotes protein synthesis (anabolic effect). Anabolic steroids have been synthesized, which have a powerful anabolic action but only a modest androgenic action. These artificial hormones are still used to produce short-term super-athletes. Such a misuse of medicine for doping purposes often results in addiction, which has serious psychological, social and physical effects. The human hypophysis produces four sex-related hormones FSH, LH, prolactin and oxytocin. LH is also called Interstitial Cell Stimulating Hormone (ICSH) in the male, because it stimulates the Leydig interstitial cells that produce testosterone, which in turn specifically inhibits LH secretion. Removal of the male pituitary causes complete loss of all testicular functions; administration of FSH and ICSH then restores these functions completely. Eicosanoids are oxygenated, unsaturated 20-carbon fatty acids that originate primarily from arachidonic acid by activation of phospholipase A2. Eicosanoids exert important effects on most human cells. Arachidonic acid is a major component of the phospholipids of membranes. Arachidonic acid is converted to prostaglandins and thromboxanes by cycloocygenases, to leucotrienes by three types of lipogenases, and to epoxides by cytochrome P-450-dependent mono-oxygenases. Prostaglandins and thromboxane (TxA2) are synthesized in response to stimuli, and they mainly act locally as autocrine or paracrine hormones. Prostaglandins (PG) are abbreviated PGD, PGE, PGF, PGG, PGH, and PGI2 (prostacyclin). Leucotrienes (LT) are abbreviated LTA, LTB, LTC, LTD, LTE, and LTF. The leucotrienes LTC4 and LTD4 are vasoconstrictors. TxA2 is not only an activator of platelet aggregation, but also an effective bronchoconstrictor, and TxA2 constricts both the cerebral and the coronary arteries. Prostaglandin E2 (PGE2) can be used to induce labour just as oxytocin. PGE2 and PGF2a increase uterine contractility by Ca2+-influx and moderation of cAMP. Prostaglandins are especially useful in second-trimester abortion. PGE is a potent vasodilatator, which can be used for intracavernous injection for impotence. Young children have low plasma [gonadotropin] from birth. The gonadotropin releasing hormone is formed in the hypothalamus (a decapeptide GnRH = LHRH), and stimulates pituitary gonadotropins to pulsate secretion of FSH and LH (= ICSH). Through childhood they develop pulsate secretion of pituitary gonadotropins, with a LH peak at night in puberty. The nocturnal LH peak disappears when adult status is reached. At the onset of puberty a timing device in the brain triggers the gonadotropin producing machinery in the hypothalamic-pituitary-testicular axis. Puberty is probably triggered by GnRH in a sufficiently mature CNS. The hypothalamic neurons mature in accordance with a genetic (familial) pattern. Puberty is a maturation process descending from the programmed brain (hypothalamus) to the pituitary gland, the gonads and eventually to the entire body. Hormones are produced at high rates, and the secondary sex characteristics then develop. Negative feedback control operates both before and after puberty, but the output of FSH and ICSH from the adenohypophysis is more than 100 times greater in young adults than in boys. Circulating inhibin is the primary inhibitor of FSH secretion by negative feedback on the pituitary gonadotropins. FSH stimulates Sertoli cells to produce more inhibin at puberty (Fig. 29-5). Circulating testosterone regulates ICSH secretion by negative feedback primarily on the eminentia mediana hypothalami. The plasma [testosterone] is highest during the night and in the morning (circadian rhythm) but there is virtually no seasonal rhythm with testosterone secretion in humans. Enlargement of the testes is the first clinical sign of male puberty. The testis consists of Leydig cells that produce testosterone. Gap junctions connect adjacent Leydig cells, and their testosterone has local nourishing effects on germ cells. The seminiferous tubules contain the germ cells (spermatogonia) and Sertoli cells. Each spermatogonium can divide into 64 spermatozoa within 65 days. The Sertoli cells secrete a wide variety of growth factors, activin, inhibin, oestrogens and an androgen-binding protein, all of which nourish the germ cells. The seminiferous tubules drain into rete testis Halleri, which communicates with the epididymis via ductuli efferentes. The epididymis is a maturation chamber for spermatozoa, where they lose their cytosol and become increasingly mobile within a few weeks. The store of mature spermatozoa is emitted into the female genitalia during copulation. The human testes of an adult male are positioned in the scrotum at a temperature around 35oC. In disease or old age the seminiferous tubules may cease functioning, but the sexual capacities (other than fertility) are well maintained as long as testosterone is produced. Aberration of sex development can arise from two different causes. 1. The sex chromosomes can create genetic sex disturbances, and 2. hormones can disturb our sex differentiation. This paragraph also deals with 3. Psychosocial sex-deviations, 4. Cryptorchism, 5. Castration, 6. Oral contraception, 7. Impotence/Prostate disorders, 8. Menstrual disorders/Occlusion of the fallopian tube, 9. Menopause, 10. Osteoporosis. 11. Breast cancer, and 12. Abortion. Sexually related infections are gathered at the end of Chapter 31. In 1938 Turner described a syndrome in small persons, retarded in growth and in sexual development. They are apparent females with small or no ovaries and a XO chromosomal karyotype. Since they have only one sex chromosome (X), their total chromosome number is 45. The Turner patient lacks the inputs from two active X chromosomes and from an Y chromosome. The lack of antimullerian hormone and testosterone leads to Mullerian duct development and female genitals, but the ovary is just a fibrous streak devoid of germ cells. The Turner patients have no sex chromatin and no drum stick (Fig. 29-6). In 1942 Klinefelter described a syndrome in persons appearing as men. These males are tall, have small dysgenetic testes, some have female breasts (gynaecomastia), and they are sterile. Their cells contain XXY chromosomes (47 instead of the normal 46). Thus Klinefelter patients must have one sex chromatin and one drumstick just like normal females (Fig. 29-6). These phenotypic XXY-males have significantly higher LH & FSH, and lower blood [testosterone] than matched XY controls. The seminiferous tubule development and spermatogenesis are deficient in Klinefelter males. The XXY-males did not show more feminine behaviour than matched controls. A similar group of tall males with XYY chromosomes were not extraordinarily masculine. Some XYY-males have significantly higher [testosterone] in their blood than matched XY controls. Some small super women have an extra X chromosome: XXX, making a total of 47 chromosomes. We expected them to have two sex chromatin and two drumsticks, and this has been confirmed. The XXX females have deficient germ cell development and often a short reproductive life.
Fig. 29-6: Intersex syndromes. Apparent men with XXXY (48) chromosomes have Klinefelter characteristics with testes, and also two sex chromatin and two drumsticks (Fig. 29-6). Individuals with four X-chromosomes are extremely rare. They are apparent females with XXXX (48), and apparent males with XXXXY (49). Cells with 4 X-chromosomes contain a maximum of 3 sex chromatin (Barr bodies) and 3 drumsticks, regardless of whether the cells come from apparent females or males (Fig. 29-6). A very small number of individuals end up being of indeterminate gonadal sex (ie, has both ovarian and testicular tissues present). Some persons have an ovary on one side and a testis on the other - a true hermaphrodite. In the Greek mythology Hermaphrodites was the child of Hermes and the beautiful Aphrodite. Pseudo‑hermaphrodites have external genitals from both sexes, but only one gonadal sex. Males have normal XY chromosomes, but small testes with poor sperms (poor spermatogenesis). Some of these genetic (XY) boys are born as apparent girls, but they may change from female to male at puberty if the penis grows. An enzyme defect that blocks the conversion of testosterone to 5-a-dihydrotestosterone disturbs the development of the external genitals. Female hermaphrodites have ovaries, female ducts, XX chromosomes, and varying degrees of masculine differentiation of the external genitals. Any XY individual with a genetic defect in testosterone synthesis develops testes due to the presence of the Y chromosome, and Mullerian duct regression due to the presence of antimullerian hormone. The Wolffian duct does not develop normally, because of the testosterone deficiency. Other XY individuals lack the androgen receptor. They develop testes (Y chromosome presence) and the so-called X-linked testicular feminisation syndrome. These XY persons show Mullerian duct regression because the antimullerian hormone is present. The lack of androgen receptors and the effects of androgens on the Wolffian ducts prevents masculinization and the external genitals are feminine. 2. Hormonal differentiation disturbances The virilising effect of testosterone on the urogenital sinus in early life causes the adrenogenital syndrome in XX individuals. They have ovaries (XX chromosome presence) and the Mullerian duct develops normally, because of the absence of antimullerian hormone. The androgen hypersecretion results in variable development of male external genitalia. The adrenal hyperplasia is caused by enzyme defects. XY individuals with deficient testosterone synthesis ability to convert testosterone to dihydrotestosterone develop testes, but the Wolffian duct structure are underdeveloped to a varying degree ranging from a partial to a complete female pattern. XY individuals who lack oestrogen receptors or have a mutant gene for aromatase, lack oestrogen effects. The functional lack of oestrogen results in unfused epiphyseal zones, so these males are tall, and they have high plasma concentrations of LH although testosterone is normal. 3. Psycho-social sex-deviations Sex identity is the individual perception of herself or himself as a female or a male. Sex identity is established early, and is not lost by castration. Both psychological and social factors can interfere with normal sexual development on the psychological plane. An imminent urge to change sex (operative sex shifts etc.) characterises trans‑sexual persons. The sex role is the social behaviour or cultural role played by or forced upon each individual. Some male homosexuals wish to express their femininity while other males clearly signal that they are men. Transvestites love to dress like the opposite sex. Transvestites are heterosexual, homosexual or asexual just as others. Cryptorchism means hidden orchids (testes). The flower orchid (French orchidé) has a root, which is actually shaped like a testis. If the testes do not descend from the abdominal cavity to the scrotum, heat destroys the sperm-producing seminiferous tubule cells. Heat does not harm the Leydig (testosterone-producing) interstitial cells. Certain cultures castrate boys to preserve their tenor voices. Puberty and natural sex development does not take place. Adult males retain their secondary sex characteristics and erection but they often lose libido. Eunuchs are more or less trustworthy in Harems. The effects of castration of adult females are surprisingly trivial, as long as the pituitary is working well. Castration, of course, stops their menstrual periodicity (artificial menopause), and they are sterile. Modern contraception is obtained with tablets (pills) containing 20-30 mg ethinyl-oestradiol and variable progesterone. The oestrogen content suppresses the hypophyseal release of gonadotropins, which prevents the maturation of the follicle, the ovulation and the luteinisation. The progesterone content favour the secretion of sperm-hostile mucus in the uterus, inhibits tuba motility and endometrial nidation. Side effects of the combined tablet are more frequent in smoking female over 35 years, and in all females with cardiovascular risk factors. Side-effects are weight gain, accentuation of cervical and breast cancer, hypertension, acute myocardial infarction, stroke, increased clotting capacity, phlebothrombosis, gallstones, hepatomes, migraine, depression, impaired glucose tolerance (Fig. 27-6), diabetes mellitus, hypercholesterolaemia, and infertility. These serious side-effects are rare but still present. Prescription of even the modern low risk pills necessitates careful control of all risk factors. Infertility is a diagnosis used on a couple, which have been unable to conceive during one year of unprotected intercourse. The causes are oligospermia, tuba blockage, ovulatory disorders, or combined problems with both persons in the couple. In some cases ovulation can be elicited by a synthetic oestrogen receptor antagonist (clomiphene), which has a high affinity towards hypothalamic oestrogen receptors. Clomifene administration simulates oestrogen deficiency in the infertile patient with a hypothalamic defect, and by negative feedback clomifene increases GnRH and FSH/LH secretion and promote fertility. 7. Impotence/Prostate disorders Impotence is frequently seen without organic cause such as hypogonadism. Diabetes mellitus, essential hypertension, and neuropathy of the autonomic system cause impotence just as antihypertensive drugs (diuretics, methylDOPA and b-blockers). Intracavernosal injections of prostaglandin E, papaverine, phentolamine, and other vasodilatating substances can provide erection for a few hours. The patient can use such a cocktail when needed. Prostate disorders, such as benigh prostatic hypertrophy and prostate cancer, increase in frequency with age above 60. Both disorders interfere with micturition and can obstruct renal function leading to renal insufficiency (Chapter 25). The enzyme 5a-reductase normally produces dihydrotestosterone from testosterone. Inhibition of this enzyme minimises the hormone conversion and causes the prostate to shrink. Prostate cancer is frequently present in males with elevated plasma concentrations of prostate-specific antigen (PSA). Manifest prostate cancer is removed immediately. In a case where surgery is containdicated, long acting GnRH agonists reduce testosterone secretion and the growth of prostate cancer. 8. Menstrual disorders/Occlusion of the fallopian tube Amenorrhoea or oligomenorrhoea are terms used for absence or irregularity of menstrual periods. Deficient GnRH release prevent FSH secretion from recruiting a dominant follicle, and complete loss of menses (amenorrhoea) may result. In oligomenorrhoea the oestrogen secretion is sufficient for uterine bleeding to occur in an irregular pattern, but often insufficient to induce a midcycle peak of LH and ovulation. Causes are ovarian disease or absence (Turners syndrome, XO), hypothalamic deficiencies, congenital adrenal hyperplasia (adrenogenital syndrome), and starvation amenorrhoea (anorexia nervosa and excessive exercise), hypothyroid amenorrhoea with increased TRH and prolactin, and withdrawal amenorrhoea (following oral contraception). Starvation amenorrhoea and anovulatory bleeding cycles often occur in female long distance runners and ballet dancers, as well as in anorexia nervosa patients (Chapter 7). These females have lost substantial amounts of fat and suffer from a serious oestrogen deficiency, which even may lead to osteoporosis (Chapter 30). Occlusion of the fallopian tube From the start of the menstrual cycle the woman is given FSH to stimulate her ovaries before ovulation. On the 12th day she is given hCG. When ovulation occurs (after 30 to 35 hours), egg cells are sucked out, placed in a tissue culture and exposed to spermatozoa. After 48 hours some eggs fertilise into the 4-8-cell stage. A few of these fertilised eggs are placed in the uterus. One in four of these eggs will nidate. Therapy is directed towards the cause of the disorder. The menopause is the event in the life of a female, where the menses stop. The last ovulations are anovulatory and conception is no longer possible. The ovaries become atrophic, the concentrations of pituitary gonadotropins (FSH more than LH) in blood plasma and in urine are the highest in the life of the female, because the follicles become more and more insensitive to gonadotropin stimulation and the oestrogen and inhibin production diminishes. Functional changes in other organs are less definitive, but vascular flushing of the head and neck are typical, probably due to the release of large amounts of hypothalamic gonadotropin releasing hormone (GnRH). Attacks of sweating during the night are classical complains. Adrenal and ovarian stromal cells secrete androgen precursors that are converted to oestrogens by aromatase in adipose tissues. This is why menopausal females with sufficient adipose tissue suffer less from oestradiol deprivation than lean females. Females with severe complains are treated with oestrogen, which ameliorates the disorders and reduces the rate of heart disease and of postmenopausal osteoporosis. Osteoporosis or thin bones is a term used for a marked reduction in all elements of bone mass. Postmenopausal females reduce their bone mass progressively with age up till the age of 70-75 years. This bone reduction also occurs in elderly males, but at a much slower rate. Elderly patients living indoors all year round are less exposed to sunshine and do not synthesise vitamin D in the skin. If their diet simultaneously is poor in vitamin D and Ca2+, it is not surprising that their bones get thin. Oestrogen therapy is beneficial as a preventive strategy in postmenopausal osteoporosis. So is increased dietary Ca2+ with vitamin D. Walking, jogging, golf are exercises retarding bone mass loss. Calcitonin has proven of benefit in some studies. A promising approach is the use of oestrogen-receptor modulators to prevent osteoporosis and thrombo-embolic events, without increasing the risk of breast cancer. Breast cancer tumours can be treated with synthetic blockers of the oestrogen receptor. The blockers suppress the growth of oestrogen-sensitive breast cancer. – Another therapy principle is to diminish oestrogen production. This is done with the drug, aminoglutethimide, which inhibits the desmolase reaction and thereby reduces adrenal steroid synthesis as a whole. Synthetic blockers of the progesterone receptor (mifepristone) induces early abortion by removing the positive progesterone effects on the conceptus. The following five statements have True/False options: A. The menarche is the last menstrual bleeding. B. Pseudo-hermaphrodites have external genitals from both sexes, but only one gonadal sex. C. HIV means Human Immunodeficiency Virus. HIV is the cause of Acquired Immune Deficiency Syndrome. HIV triggers a progressive and irreversible depletion of T-helper lymphocytes. D. Transvestites love to dress like the opposite sex. Transvestites are heterosexual, homosexual or asexual. E. At the onset of puberty a timing device in the brain triggers the Gonadotropin producing machinery in the hypothalamic-pituitary axis. A 24-year-old female is going through her last menstrual cycle before pregnancy. 1. Summarise schematically the most important hormonal events in her menstrual cycle. 2. Summarise schematically the most important hormonal events during continued pregnancy and delivery. A pregnant woman delivers oxygen to her foetus. Her A‑haemoglobin (A = adult) is functionally different from that of her foetus (F‑haemoglobin). 1. Why is this difference important? How are the two dissociation curves related? 2. FSH and LH are important for this woman. Describe why. Describe the function of the two hormones in her husband. 3. Following birth the mother breastfeed her baby and experience a feeling of sexual pleasure including uterine contractions. Describe the mechanism. See answers · The presence of normal ovaries or testes determines the gonadal sex. Without normal ovaries or testes any genetic sex will develop into an apparent female. · The brain is an important sex organ. The sex desire (libido) is stimulated by a multitude of sense impressions (visual, auditive, olfactory, and psychological). Potency refers to the ability to engage in intercourse. · On the first day of the menstrual bleeding, the low progesterone and high prostaglandin level probably releases enough Ca2+ to start spontaneous contractions of the myometrial cells. Ca2+ -ions enters myometrial cells and stimulates their activity in the secretory (progesterone) phase. · At certain high plasma level of oestradiol can increase FSH output. This is called the positive feedback release ovulation. At lower levels oestradiol is a potent inhibitor of Gonadotropin-RH secretion and thus of FSH output (negative feedback). The negative feedback forms the basis for the ovulation-inhibition by contraceptives. · The primary inhibitor of FSH secretion is the peptide, inhibin that is secreted by the ovary and testis, and blocks the effect of Gonadotropin-RH. · The plasma [oestradiol] increases sharply in the last part of the follicular phase, while the [LH] also increases. The sharp rise in LH and a modest rise in FSH coincide with ovulation. The LH not only causes rupture of the follicle; it continues to act on the follicular cells, turning them into a yellow endocrine organ, the corpus luteum. · The spermatozoa can keep their vitality for more than 4 days if they reach the tube. They lose their protection cover in the uterine tube. The head of the spermatozoa swell and liberates proteolytic enzymes. These enzymes dissolve the corona radiata around the egg (oocyte). The oocyte can only live 14 hours without conception. · Due to the priming effect of oestrogen on progesterone receptors, both hormones stimulate the growth of the endometrial glands, so that they curl like a helix. The progesterone effect in particular provides the endometrial/myometrial tissues with their high secretion and blood perfusion, so the uterus is prepared to receive the fertilised ovum. · The b-group of hCG is specific and found in the blood by specific antibody methods even before the first menstrual bleeding fails to appear. The hCG is detectable in the urine 8-12 days after the first missing vaginal bleeding. · During puberty FSH, LH, growth hormone, and insulin are important for the breast development. The thyroid hormones (T3/T4) are permissive. At the end of pregnancy there are other hormonal events. Progesterone secretion reaches a peak and then falls. This fall in progesterone allows the pituitary to release prolactin (LTH). · Relaxin is a pro-insulin-like polypeptide produced by the corpus luteum. The hormone relaxes pelvis articulations and softens the uterine cervix in order to facilitate passage of the foetus. These and several other factors are involved in human labour, but the exact trigger mechanism remains unclear. · Turner described a syndrome in small apparent females, retarded in growth and in sexual development, and with small or no ovaries. Since they have only one sex chromosome (X), their total chromosome number is 45. They have no sex chromatin and no drumstick. · Klinefelter described a syndrome in persons appearing as males. They are tall, have small testes, some have female breasts (gynaecomastia), and they are sterile. Their cells contain XXY chromosomes (47 instead of the normal 46). · Amenorrhoea or oligomenorrhoea are terms used for absence or irregularity of menstrual periods. Causes are ovarian disease or absence (Turners syndrome, XO), hypothalamic deficiencies, congenital adrenal hyperplasia (adrenogenital syndrome), starvation amenorrhoea such as in anorexia nervosa and excessive exercise, hypothyroid amenorrhoea with increased TRH, and withdrawal amenorrhoea (following oral contraception). Johnson MH and BJ Everett. Essential reproduction. 6th Ed. Blackwell Science, Oxford, 2007.
|
||

.jpg){kind=link}
{kind=link}
{kind=link}
Click here to introduce your comments or contributions