New Human Physiology | Paulev-Zubieta 2nd Edition
Chapter 3: The Somatosensory System And Disorders
| HOME | PREFACE | TABLE OF CONTENTS | SYMBOLS | SECTION INFO | CONTRIBUTORS | LINKS | CONTACT US |
Highlights
Study_ObjectivesPrinciplesDefinitionsEssentials
PathophysiologyEquationsSelf-AssessmentAnswers
Further Reading
|
Chapter 3
|
 |
|||||||||||||||||||||||||||||||||||||||||||||
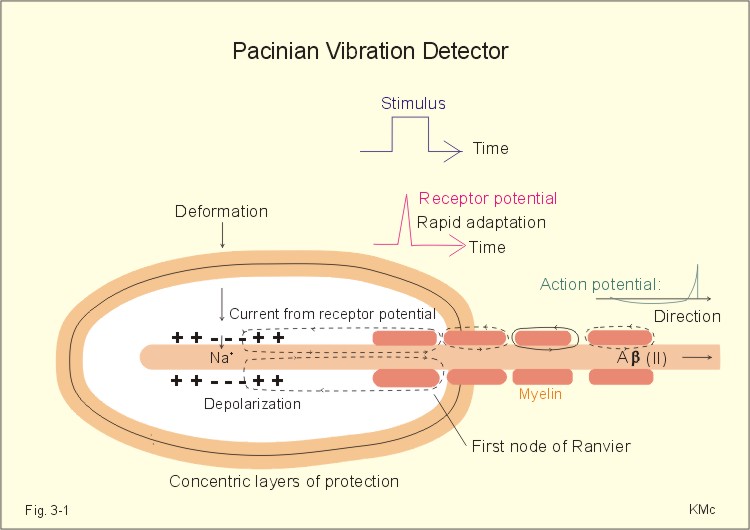
Adaptation or accommodation of sensory receptors refers to a progressive decrease in firing frequency despite maintained depolarisation. Adequate stimulus refers to the stimulus, for which the receptor has a lower energy threshold than for other stimuli - ie, the stimulus to which the receptor is most sensitive. AMPA is an abbreviation of alpha-amino-3-hydroxy-5-methyl-4-isoxazole-propionic acid. This is an excitatory amino acid (EAA) and one of the EAA-gated ion channels carries AMPA-receptors with high affinity for a subclass of glutamate receptors. Causalgia means hyperalgesia (hypersensitivity to pain) elicited by a cold stimulus. Coding: In neurons the stimulus intensity is coded by the frequency of action potentials. Dermatomes: Every spinal dorsal root is destined to a segment of the skin called a dermatome. Endogenous opioids are substances in the CNS with opiate-like effects. GABA is the abbreviation for gamma-amino butyric acid – the most common inhibitory transmitter in the brain. The gate-control hypothesis of pain states that pain transmission is suppressed by innocious signals in thick myelinated afferents (group II), whereas the pain sensation is enhanced by signals in thin afferents. Headache is pain on the surface of the head, which is actually due to anomalies in intracranial or extracranial structures. Hyperalgesia means hypersensitivity to pain. NMDA is the abbreviation for N-methyl-D-aspartate. Nociceptors or nocireceptors are responsive to stimuli that potentially cause injury. Plasma membrane receptors consist of a protein or glycoprotein molecule, an ion channel pore or a specific enzyme (G-protein). Receptor potential: The stimulation of a receptor elicits a generator potential that is graded according to the stimulus strength. When the stimulus is strong enough to reach the threshold, an action potential is fired. Sensory receptors translate stimulus intensity to impulse activity in sensory afferents. Sensory receptors are either neurons in the case of vision, smell and cutaneous senses, or modified epithelial cells in the case of auditory, vestibular and taste senses. Threshold stimulus refers to the weakest stimulus to which the receptor will react. Trigeminal neuralgia (French: Tic douloureux) is a condition with daily paroxysms of violent pain in part of the trigeminal area lasting only some seconds. The pain is provoked by eating, washing the face or by cold in the face. This paragraph deals with 1. Sensory receptors and nerve fibres, 2. Blood-brain barrier and CSF, 3. Regeneration of nervous tissue, 4. Sensory pathways, 5. Central opiate receptors, 6. Taste and smell. 1. Sensory receptors and nerve fibres Sensory receptors are either neurons in the case of vision, smell and cutaneous senses, or modified epithelial cells in the case of vision, auditory, vestibular, smell and taste senses. The special sensory receptors for vision, hearing and balance are described in Chapter 5. Some sensory receptors have characteristics similar to the well-known plasma membrane receptors. Plasma membrane receptors consist of a protein or glycoprotein molecule, an ion channel or a specific enzyme (G-protein). The stimulation of a receptor elicits a receptor potential (generator potential) that is graded continuously with stimulus intensity. When the stimulus is strong enough to reach the threshold, action potentials (APs) are fired. In neurons the stimulus intensity is coded by the frequency of action potentials. Sensory receptor systems are biological transducers with a dynamic range of up to 1012 in the most sensitive organ, the ear. The threshold is the reciprocal of the sensitivity. The threshold is the weakest stimulus to which the receptor will react. The sensitivity of a sensory receptor is greater the smaller its threshold stimulus is. The Pacinian corpuscles in the skin are gigantic receptors (almost 1 mm long) consisting of concentric layers like onion-scales in the microscope. There is a single axon in the axis of each corpuscle (Fig. 3-1). Stimulus intensity is coded by the single axon. Compression deforms the axon and depolarises the membrane by opening of Na+-channels.
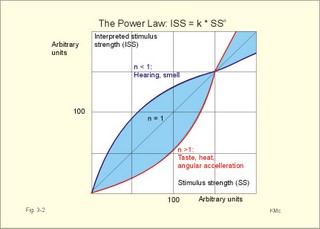
Fig. 3-1: The Pacinian corpuscle (1 mm long) is a vibration detector. The depolarisation generates a graded receptor potential forcing a current towards the first node of Ranvier with maintained stimulus. The receptor potential rapidly decreases (rapid adaptation), because the adequate stimulus is alterations in the deformity rate (vibrations in the range 150-300 Hz). At the first node of Ranvier, a propagating action potential along the axon is released, provided the generator potential is sufficiently large (Fig. 3-1). The Pacinian corpuscle is located in the deeper layers of the skin and connective tissue. The afferent fibres are thick (Type Ab or II), and they lead the signals to synapses in nucleus gracilis and cuneatus of the spinal cord. Steven proposed the power law that is given as Eq. 3-1. The interpreted stimulus strength (ISS) is equal to a constant (k) multiplied by the actual stimulus strength (SS) raised to the power n. The situation n equal to 1 describes a linear relation between stimulus and resulting activity in the conducting neuron (Fig. 3-2). Perception of taste, heat and angular acceleration are described by power functions or transfer functions with n just above 1, whereas hearing and smell are described by functions with n lower than 1. The only sensation with a particularly large value of n (about 3) is pain, which is why pain is felt so severe with increasing stimulus intensity!
Fig. 3-2: A graphical description of the power law. However, the power law and other types of curve fitting with transfer functions have hardly improved our understanding of sensory modalities. Both conduction velocity and size is used in classification of nerve fibres. The fibres are divided into types A, B and C, based on the three main conduction velocities shown in the record of the compound action potential from a mixed nerve. Type A, B and C refer to phases in the combined action potential. Type A fibres are the fast conducting myelinated fibres (thick fibres subdivided into a , b , g , and d ), type B are preganglionic sympathetic fibres, and type C are the small, unmyelinated fibres. Another classification is based on the thickness of the axons (I-IV). The size classification became necessary, when Aa - fibres were separated into two subgroups: Ia and Ib. In Table 3-1, the velocity classification (A-C) is written first, and the size classification (I-IV) is given in parentheses. _______________________________________________________________________
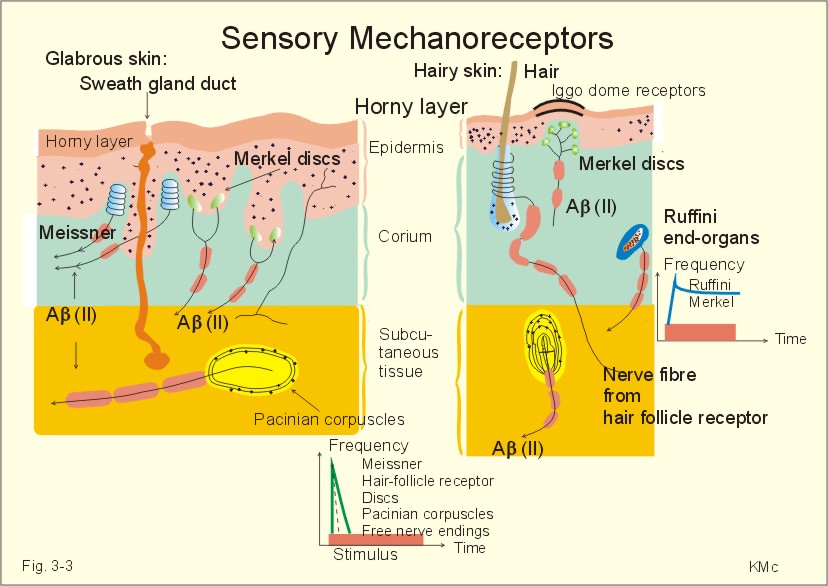
The Aa (I) fibres are motor a -fibres and proprioceptors from the annulospiral endings of muscle spindles (Ia) and from Golgi tendon organs (Ib). The Ab (II) fibres conduct discrete touch and fine pressure signals from cutaneous tactile receptors. The Ag (II) fibres are motor fibres to muscle spindles. They have their origin in the spinal cord. The Ad (III) fibres transfer pain sensations, decline in skin temperature as well as crude, passive touch and deep pressure. The B fibres (III) are autonomic preganglionic fibres. The C fibres (IV) are unmyelinated and lead pain, touch and signals from heat receptors from the skin. The C fibres have no myelin sheath. Sensory receptors in the nervous system are classified as exteroceptors (located on the body surface), proprioceptors (located in muscles, tendons and joint capsules), interoceptors (located in the viscera), and telereceptors (stimulated by events far from the person). Cutaneous receptors are exteroceptors (Fig. 3-3). Pacinian and Meissner corpuscles are rapidly adapting (dynamic) touch velocity detectors in glabrous skin. In hairy skin, hair-follicle receptors are velocity detectors (they adapt rapidly). Meissner corpuscles are located in the papillae of the hairless skin such as fingertips, lips and clitoris. Merkels discs and Ruffini-end organs are slowly adapting (static) touch intensity detectors both in hairless and hairy skin. Merkels discs are found in elevated dome corpuscles in hairy skin (up to 50 Merkel discs in a corpuscle of 0.5 mm in diameter). These so-called Iggo dome receptors are extremely sensitive and transmit touch signals to a single nerve fibre. In this way, even weak tactile stimuli can create sensation in the CNS.
Fig. 3-3: Cross section through an area of hairless and a hairy skin showing 6 types of sensory mechanoreceptors. All the mechanoreceptors are supplied by afferent nerve fibres of group II, except the free nerve endings (group IV). The free nerve endings, with unmyelinated afferents (group IV) conduct signals with low velocity (Table 3-1), and register passive touch, such as, slow strokes with a piece of cotton. Thermoreceptors are also exteroreceptors. We have cold-receptors just below the skin surface (200 nm deep). Cold receptors respond to changes in temperature. Heat receptors are also located in the skin. The location of certain heat and cold points in the skin is determined by bringing a thin hot or cold object in contact with the skin. Thermoreceptors react to temperature changes. Cold receptors and heat receptors in the skin are located close to the surface. Both types of receptors are also located in the deep tissue and in the CNS. Both types of receptors discharge spontaneously at normal temperature, dynamically when skin temperature is changing rapidly, and adapt slowly. Proprioreceptors, located in muscles, joints and joint capsules, are mechanoreceptors (muscle spindles, Golgi-receptors, Pacinian and Ruffini corpuscles, and free nerve endings). The Ruffini mechanoreceptors are also called joint receptors, because they are located in ligaments, tendons and articular capsules. They provide information for the CNS concerning articular movements, movement velocity and joint position. Joint receptors of the proximal joints are particularly sensitive. The static and dynamic receptors inform the CNS about the position and movement of the joint, respectively. These receptors enable us to sense the position of the joint with great accuracy. Accommodation of sensory receptors or adaptation is a progressive decrease in firing frequency despite maintained depolarization. The frequency of action potentials from stimulated receptors fall, although the stimulus is maintained at constant strength. Accommodation or adaptation occurs, when a proportion of the voltage-gated Na+-channels is rapidly inactivated by depolarisation, which also opens K+- channels. This makes the cell more refractory to stimulation. Accommodation can also be caused by a hyperpolarization induced by gradual activation of Ca2+ -dependent K+ -channels.
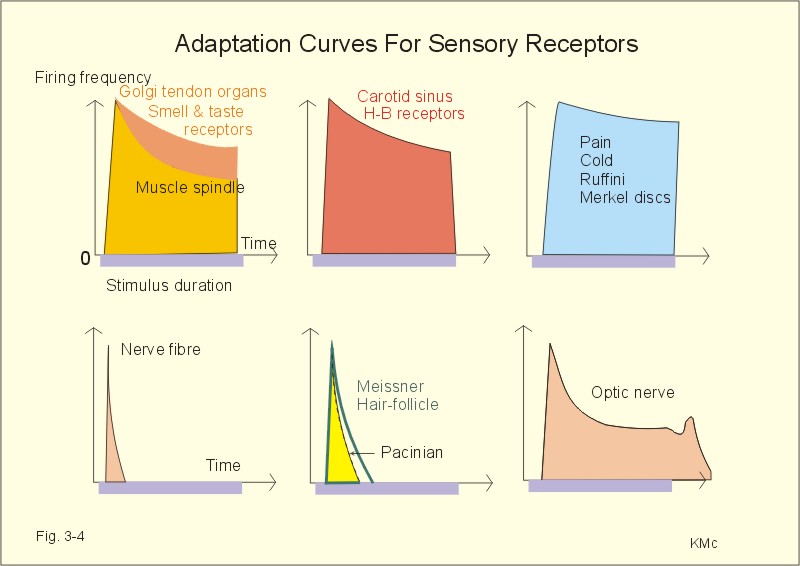
Fig 3-4: Accommodation or adaptation curves from different sensory receptors. Pain- and cold-receptors, Merkel discs and Ruffini-end organs adapt extremely slowly and incompletely (Fig. 3-4). Joint receptors, smell-taste-receptors, muscle spindles, carotid sinus- and pulmonary stretch receptors, and the optic nerve, all adapt somewhat better (Fig. 3-4). Hair-follicle receptors, Meissner corpuscles and Pacinian corpuscles adapt rapidly, just as many free nerve endings (Fig. 3-4). Nociceptors or nocireceptors (pain receptors) are responsive to stimuli that potentially cause injury. Nociceptors are free nerve endings of two types. The fast adapting Ad fibre mechanical nociceptors (group III) are high-threshold, finely myelinated afferents that originate superficially in the skin. The slowly adapting C-polymodal nociceptors (group IV) are unmyelinated afferent fibres that originate in the deeper cutaneous tissue, and respond to various mechanical, thermal and chemical stimuli (Fig. 3-5). In the spinal cord nociceptive afferents synapse with secondary neurons in lamina I and II. These sensory neurons ascent in the spinothalamic tracts. The fast adapting pain through group III fibres is bearable (acute, sharp, stinging, somatic pain), compared to the slowly adapting unbearable pain (diffuse, burning, prolonged secondary, visceral pain) through group IV fibres.
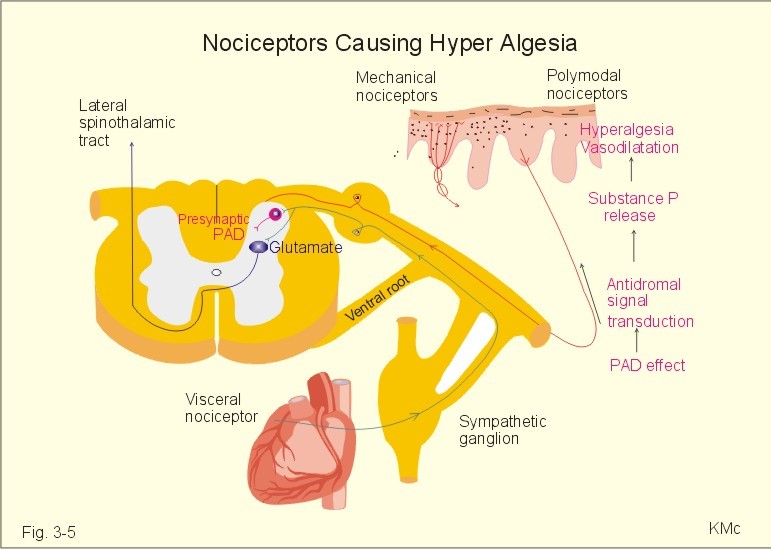
Fig 3-5: Mechanical, polymodal and visceral nociceptors. A visceral pain afferent synapses in the spinal cord with the neuron of the lateral spinothalamic tract on which the cutaneous group IV pain afferent terminates. When nociceptors become sensitised (ie, more responsive), their thresholds are reduced, thus causing hyperalgesia (ie, hypersensitivity to pain). Many substances such as bradykinin, histamine, leucotrienes, prostaglandins, serotonin, and K+ that are often released near damaged or dying cells sensitise nociceptors. K+ activates the nociceptors. Substance P is also released from polymodal nociceptors through an axon reflex with antidromal signal transduction in afferent group IV fibres, causing hyperalgesia, vasodilatation and increased capillary permeability (Fig. 3-5). Glutamate may be co-released with substance P from the polymodal C-fibre terminals. The gate-control hypothesis of pain states that pain transmission is suppressed by innocious signals in thick myelinated afferents (group II), whereas the pain sensation is enhanced by signals in thin afferents. Inhibitory interneurons of the lamina II in the dorsal horn of the spinal cord perform the gate-control through a special type of presynaptic inhibition called primary afferent depolarization (PAD), and the receptors on the cell body of the secondary neuron is the gate. The gate control hypothesis explains why innocuous signals, mediated by large myelinated afferents, can inhibit pain mediated by thin myelinated afferents. The adequate stimulus is the stimulus, for which the receptor has a lower energy threshold than for other stimuli (ie, the stimulus to which the receptor is most sensitive). The adequate stimulus for pain receptors is mechanical deformation, extreme temperature or tissue damage. The sense impression depends on the site in the brain which receives the sensory signal (ie, central analysis) and on the receptor localisation (ie, peripheral analysis). This is how different neurons transmit different types of sensations, even though they may transmit the same electrical signals (see Chapter 8). The CNS discards more than 99% of all incoming signals as irrelevant. The visual system is an example of a specific information line for a certain modality of sensation. The neurons in the retina, the optic nerve, the lateral geniculate nucleus, and the visual cortex describe just such a dedicated neuronal pathway. The specific information line through which the signal is conducted determines the way in which a suprathreshold stimulus is perceived (eg, pressure applied on the eye will be perceived as light). The auditory system also forms a specific or labelled line all the way from receptor to cortex. In all cases the specific region in the cerebral cortex, where the nerve fibre ends determines the modality of sensation. Now, where is the sense interpretation localised and what is its intensity?
2. Blood-brain barrier and CSF The blood-brain barrier consists of tight junctions between the endothelial cells of the capillaries in the CNS and of neuroglia. This barrier only allows extremely small or hydrophobic molecules to pass into the brain. The cerebral microcirculation consists of strong arterioles that can constrict to carry a high arterial pressure without brain oedema. Many large molecules cannot pass from the blood to the cerebrospinal fluid (CSF) across the choroid plexus, a tight junction barrier that is called the blood-cerebrospinal fluid barrier. The blood-CSF barrier of the choroid plexus allows some large molecules to pass from the blood to the CSF.
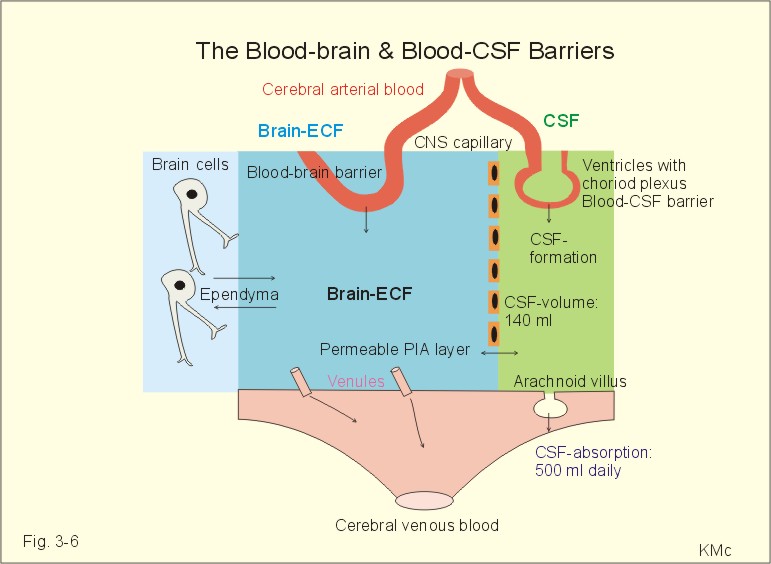
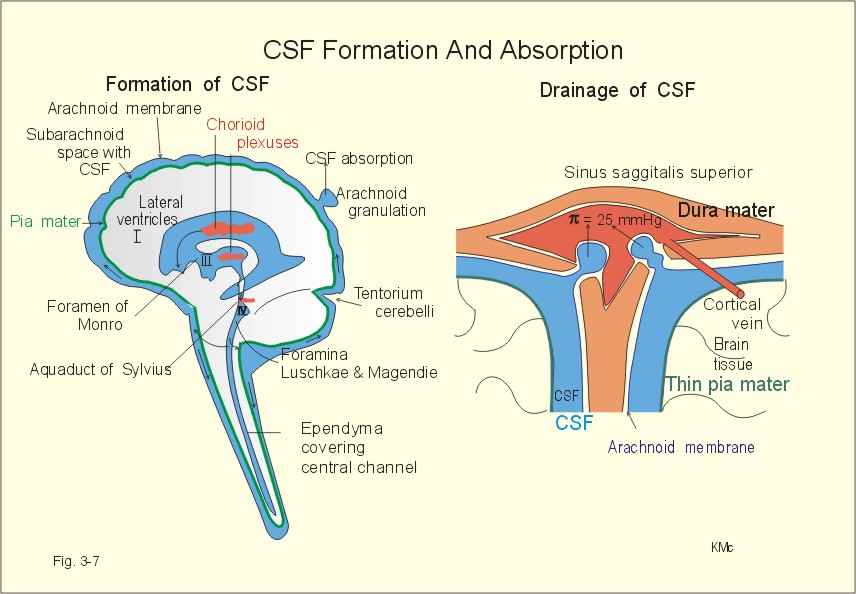
Fig. 3-6: The blood-brain and blood-CSF barriers showing the daily formation of 500 ml of CSF. The blood-brain and the blood-CSF barriers exist in all areas of the brain, except in the so-called circumventricular organs (hypothalamus, the pineal gland, and the area postrema). These discrete organs have highly fenestrated capillaries that are easily penetrated by large and small molecules as well as ions. The circumventricular organs are located close to essential control centres in the hypothalamus and brain stem regions regulating respiration, blood glucose concentration, and extracellular fluid osmolality. The two brain barriers are almost impermeable to large molecules such as plasma proteins, but highly permeable to CO2, oxygen, water, alcohol, anaesthetics, hallucinogens, and other lipophilic substances. The blood-brain barrier is almost completely impermeable to water-soluble molecules, electrolytes such as H+, whereas CO2 passes through the barrier to the medullary chemoreceptors (Fig.16-3). Humans produce 500 ml of CSF daily. The total CSF volume is only 1/3 of the daily production. Most of the 500 ml of CSF is produced in the choroid plexuses in the four brain ventricles, and the remaining is produced across the blood-brain barrier. The ventricular system and the central spinal channel are covered with ependyma. The absorption of CSF takes place through the arachnoidal granulations, which protrude, into the sinus sagittalis. The rate of absorption is directly related to the pressure in the cranial cavity - in particular the CSF- pressure. Proteins can pass through large holes in the endothelial cells. The CSF is separated from the brain cells by the thin pia mater. Substances that enter the CSF can easily diffuse into the brain interstitial fluid. Drugs that cannot pass the blood-brain barrier can enter the brain through pia mater, when infused into the CSF (Fig. 3-6). The CSF passes from the lateral ventricles (I and II) through the foramen of Monroe into the third ventricle (III), through the aquaduct of Sylvius, the fourth ventricle (IV), and out into the subarachnoid space through the foramina Luschkae & Magendie (Fig. 3-7).
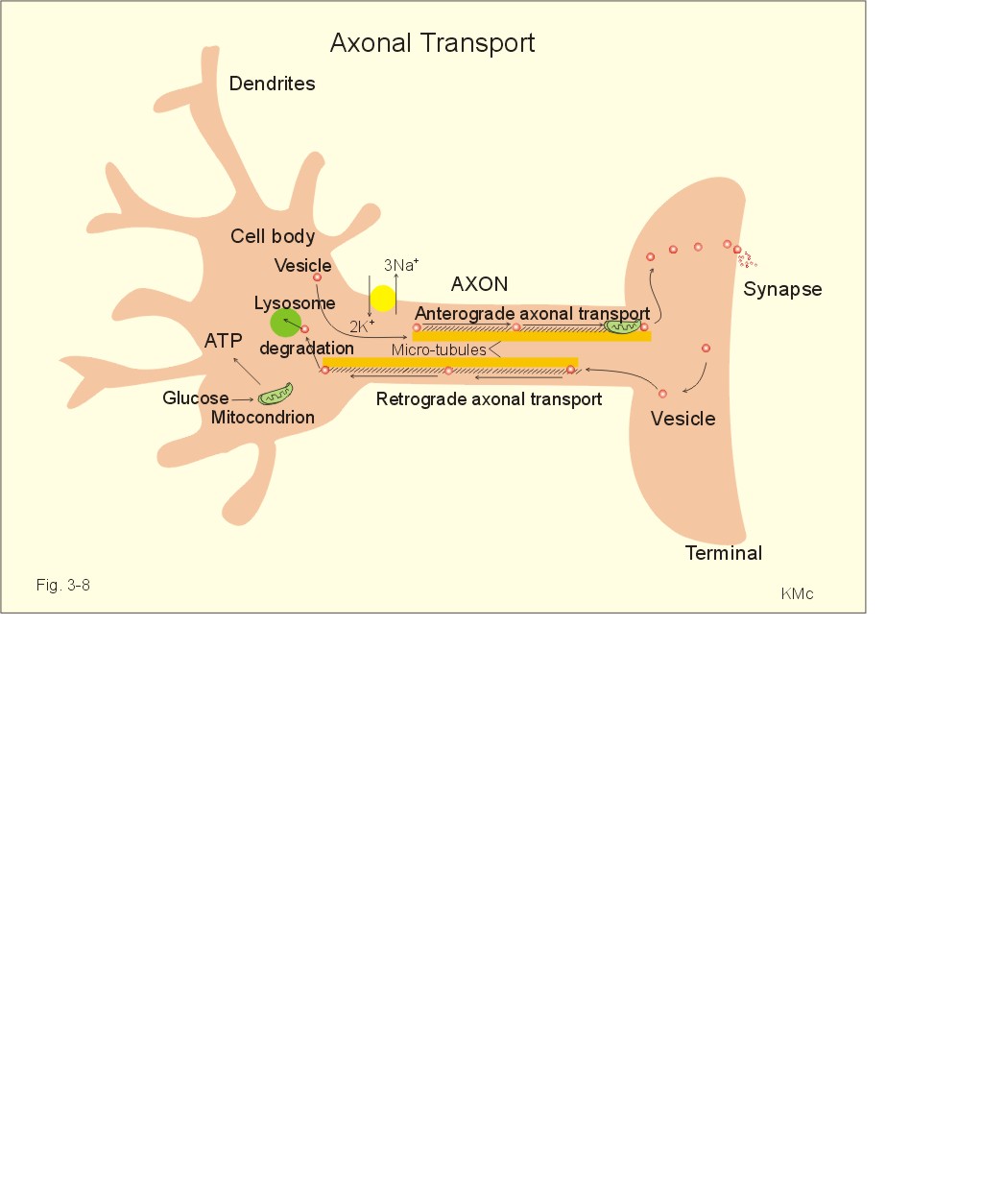
Fig. 3-7: Anatomical structures involved in CSF-formation and absorption. The normal CSF-pressure in a supine person is up to 10 mmHg (1.3 kPa) or 136 mm of water. The secretion of fluid by the choroid plexus depends on the active Na+-transport across the cells into the CSF. The electrical gradient pulls along Cl-, and both ions drag water by osmosis. The CSF has lower [K+], [glucose], and much lower [protein] than blood plasma, and higher concentrations of Na+ and Cl-. The production of CSF in the choroid plexuses is an active secretory process, and not directly dependent on the arterial blood pressure. The CSF is separated from the brain cells by the extremely thin pia mater. All natural substances that enter the CSF can easily diffuse into the brain extracellular fluid (Fig. 3-6). CSF leaves the four ventricles through the roof of the 4th ventricle, traverses the subarachnoid space, and is reabsorbed into the blood of the venous sinuses via the arachnoidal villi. The absorption here is directly related to the CSF pressure in the cranial cavity. Large holes through the endothelial cells allow proteins to enter the blood. 3. Regeneration of nervous tissue Severe injury to nervous tissue causes cell death. Neurons are postmitotic cells. For this reason lost neurons cannot be replaced. There is, however, considerable capacity for regeneration of axons in the peripheral nervous system. Both growth and maintenance of axons require the nerve growth factors (NGF). NGF is an essential survival factor for neurons outside the CNS - in particular sensory neurons. NFG binds to receptors belonging to the insulin receptor family (tyrosine kinase family). When a motor axon has been severed, the cell body undergoes chromatolysis. This is a neuronal reaction, where the rough endoplasmic reticulum (the Nissl bodies) becomes active. The Nissl bodies accumulate proteins required for repair of the axon. The axonal reaction is an attempt to repair the fibre by production of new protein structures that are transported along the axon. Therefore, proteins distend the rough endoplasmic reticulum. The axon and the myelin sheath distal to the injury die and are phagocytized. The neuroglial Schwann cells that had formed the myelin remain alive. This is the so-called wallerian degeneration named after Waller. The Schwann cells proliferate and form long rows along the pathway previously occupied by the dead axon. The severed axon regenerates along this pathway, and growth cones may eventually reinnervate the target organ. Neurological injury probably involves excessive glutamate receptor stimulation as a common pathway. Glutamate is the most important of the excitatory amino acids (EAAs) in the spinal cord and the brain. Glutamate stimulates the family of EAA-receptors including AMPA-, NMDA-and metabotropic receptors. NMDA means N-methyl-D-aspartate. - Effective glutamate antagonists are applied in clinical studies of pain. The inhibitory amino acids, GABA and glycine, and the monamines and endogenous opiods inhibit the second-order neurons of the spinothalamic tract. Fast axonal transport of organelles in the cytosol occurs as rapidly as 0.4 m per day. At this rate synaptic vesicles can travel along the motor axon from the spinal cord to a patient's foot within three days. Fast axonal transport of enzymes and organelles occurs on microtubuli in the axons, and is not interrupted by resting periods in cell compartments outside the transport system (Fig. 3-8). Oxidation of glucose in the mitochondria provides ATP for the Na+-K+-pump and for transport filaments and microtubules embedded in the axonal cytoplasm (Fig. 3-8).
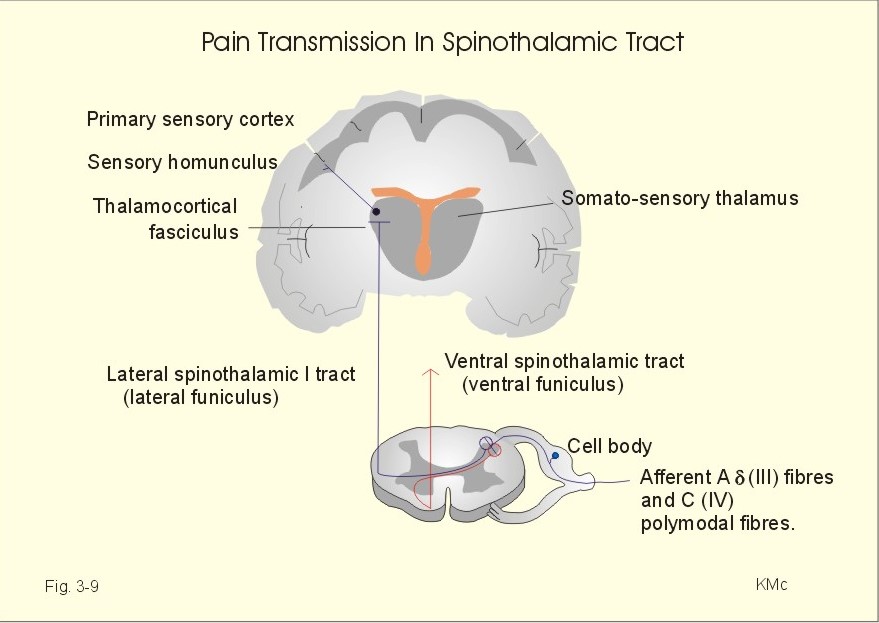
Fig. 3-8: Axonal transport of vesicles, organelles and proteins by microtubuli. Slow axonal transport occurs as diffusion of cytosolic proteins and organelles such as mitochondria. This transport occurs at a rate 100 times more slowly than fast axonal transport. Organelles or enzymes are stored in different cell compartments on their way or their direction of transport reverses. Axonal transport can be anterograde, when it occurs in the direction from the soma to the axonal terminals. Axonal transport can also be retrograde, when it occurs in the opposite direction. Here vesicles are degraded by lysosomes, when returned to the soma. A typical example of slow transport is the transfer of the many mitochondria towards the terminal of an axon. In the CNS, fast neurotransmission is inhibitory or excitatory. In the neuromuscular junction, each signal is always excitatory and sufficient to trigger a muscular contraction. In the neuromuscular junction, acetylcholine is the only neurotransmitter, whereas in the CNS there is a large variety of neurotransmitters (see Table 7-1 and 7-2). The sensory system transmits signals from sensory nerve receptors in the body. The nerve receptors are located in the skin, muscles, tendons, joints and viscera. The signals are transferred to the CNS by a pathway of first, second, third, and higher-order neurons. The third and higher order neurons are located in the thalamus and the cortex. The cell body of the first order afferent neuron is located in the dorsal root or in the cranial nerve ganglia. The signals pass through the spinal cord, the brain stem, and the thalamus before reaching the cerebral cortex. Several sensory tracts and pathways synapse in the nuclei of the thalamus (the spinothalamic tracts). The somatosensory thalamus is a relay station for most sensory modalities. The sensory inputs are processed in somatotopic areas of the thalamus, and are then transferred to appropriate cortical areas. The somatotopic organisation is maintained all the way to the cortex. The reticular activating system (RAS) of the brainstem is involved in arousal acting in concert with the thalamus. The spinothalamic tract conveys pain and temperature (lateral tract), and also crude passive touch (ventral tract). The first-order neurons are afferent Ad fibres (III) which have cell bodies in the spinal ganglia. Second-order neurons cross immediately to the opposite side of the spinal cord, and ascend in the lateral and ventral spinothalamic tract.
Fig. 3-9: The spinothalamic tracts and their sensory function. Pain and temperature reach the thalamus in the lateral spinothalamic tract (in the lateral funiculus). The second-order axon terminates in the somatosensory thalamus (the ventral posterior lateral nucleus and the central lateral nucleus). The third-order neurons pass from the somatosensory thalamus via the thalamocortical fasciculus to the somatosensory cortex or the primary sensory cortex (somatic sensory area I, or area 1, 2, 3 in Fig. 4-2) with the sensory homunculus. Some third-order neurons also pass to the somatic sensory area II of both hemispheres. Proprioception and active tactile signals are transmitted through sensory nerve fibres to the spinal cord. Primary afferent fibres ascend in the dorsal columns all the way to the medulla oblongata. These primary axons synapse with second-order neurons in the gracile and the cuneate nuclei. These second order neurons cross the midline in the medulla, and ascend in the medial lemniscus to end in the somatosensory thalamus. The medial lemniscus pathway transmits proprioception and fine tactile senses. The spinothalamic tract is the most important pathway for pain. The second order neurons of the spinal tracts have their cell bodies in the lamina I, II and V of the spinal cord. These cells receive excitatory signals from nociceptors in the skin, muscles and viscera. The action potentials from the nociceptors are conducted along the axon to the spinal cord and release neurotransmitters such as the excitatory amino acid, glutamate, and different neuropeptides. When these neurotransmitters bind to the receptors on the postsynaptic membrane of the secondary neurone, they increase the permeability to small ions, and excite secondary, postsynaptic neurons. The secondary neurons of the spinothalamic tract projects mainly to the contralateral thalamus by crossing over immediately through the anterior commissure to the opposite side of the spinal cord within the incoming segment. The endogenous analgesia system is a pain control system descending from brainstem to the spinal cord (Fig. 3-10). As an example, this system may explain why a runner who twists his leg during a competition may finish the run before he really feels the pain. As soon as he has passed the goal and stop running the pain often becomes severe, and he cannot run at all. The cell bodies of the neurons belonging to this system are located in the periaqueductal grey area of the midbrain, the periventricular areas, locus coeruleus, and the areas surrounding the aqueduct of Sylvius (Fig. 3-10). Signals from these cell bodies reach the medullary raphe magnus nucleus and the medullary nucleus reticularis gigantocellularis with nucleus reticularis paragigantocellularis lateralis. The nuclei transmit signals via the descending pain-suppressing pathway in the dorsolateral column to a pain inhibitory complex. Stimulation or increased tone of the analgesia system can suppress strong pain signals entering the spinal cord through the dorsal spinal horn. These regions contain opioid receptors. There are at least 4 types of central opiate receptors and their subtypes: u for morphine-like drugs, d and k for enkephalins, and the non-selective s -receptors. Endogenous opioids are substances with opiate-like effects. These substances are naturally occurring in the nervous system (b -endorphin, met-enkephalin, leu-enkephalin, dynorphin and many others). Endogenous opioids are derivatives of three large protein molecules encoded by three different genes. These mother-molecules are pro-opio-melanocortin (POMC), proenkephalin and prodynorphin. 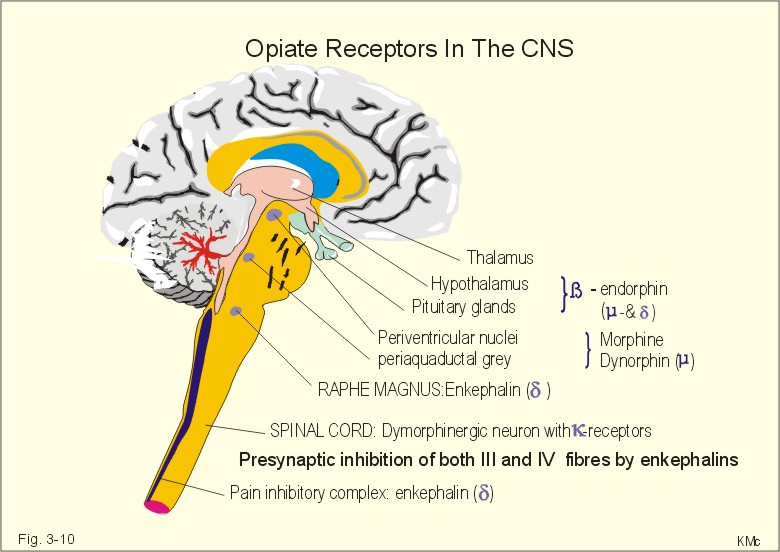
Fig. 3-10: Opiate receptors in the CNS. Enkephalin, b-endorphin and dynorphin (3 peptides) appear in normal human cerebrospinal fluid. Enkephalins inhibit both type C and type Ad (III) pain fibres presynaptically in the dorsal horns. Enkephalin is the endogenous ligand for the d -opiate receptors. Dynorphin has much higher affinity than morphine and is only found in small quantities close to the dynorphinergic k -opiate receptors. b -endorphin is present in the hypothalamo-hypophysary system. Presynaptically located opiates inhibit depolarization of nerve terminals and reduce synaptic transmission. The purpose of pain is to protect the body from further or imminent harm. A special type of burning pain is provoked by noxious heat or by capsaicin (which contain a vanillyl-group) in chilli, paprica and pepper. These spices and heat stimuli seem to activate a vanniloid receptor subtype 1 in sensory nociceptors with terminals in the dorsal horn of the spinal tract. Activation opens Ca2+-channels and the Ca2+-influx is probably involved in the burning sensation. Gyrus cinguli has the highest density of central opiate receptors. Pyramidal cells are contacted by opiate secreting interneurons that inhibit arriving pain signals. The sensations from the anterior 2/3 of the tongue travel with the trigeminal nerve fibres, through the chorda tympani into the facial nerve (VIIth), and eventually reach the solitary tract of the brain stem. Taste signals from the back of the tongue and surrounding tissues are transmitted through the glossopharyngeal nerve (IXth) into the tractus solitarius. All taste fibres synapse in the nuclei of the solitary tract and the axons of these neurons project to the thalamus. From the thalamus third-order neurons reach the lower part of the primary sensory cortex in the postcentral gyrus (somatosensory area I = area 1 in Fig. 4-2).
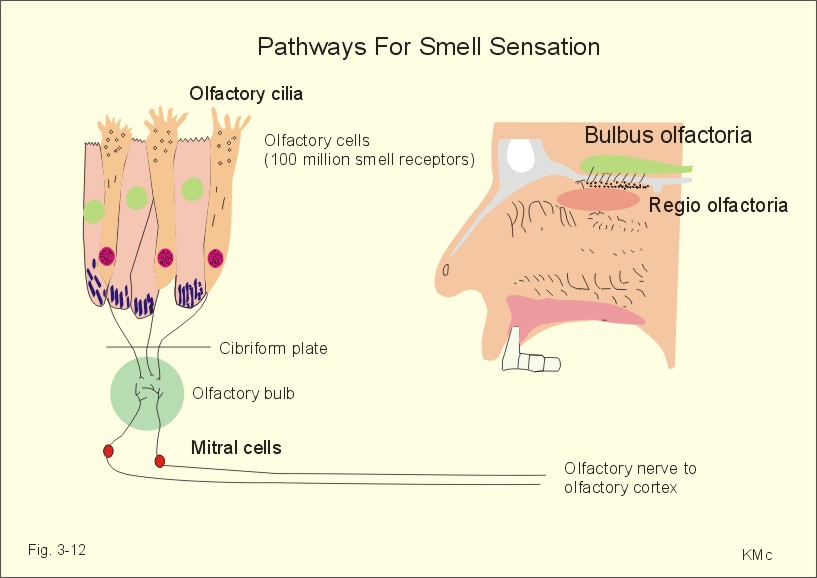
Fig. 3-11: Taste buds and taste pathways from the tongue. Acids evoke sourness, because H+ stimulates special H+-receptors in the taste buds. Saltiness is produced by the anions of inorganic salts. The Cl--receptor is particularly effective in registering saltiness. Our taste buds at the base of the tongue also have bitter-receptors stimulated by many long-chain organic compounds. Many alkaloids (quinine, caffeine, and nicotine) also taste bitter. Sweet-receptors are stimulated by sucrose, glucose, lactose, maltose, glycerol, alcohol, aldehyde, ketone, and organic chemicals. In the upper nasal cavity the mucous membrane is yellow and termed the olfactory membrane. It contains 100 million bipolar neurons called olfactory cells. They contain hairs or olfactory cilia. The olfactory cells are smell receptors. They work as telereceptors, and the smell pathways do not include the thalamic relay station and a neocortical projection area. Instead, the olfactory cells pierce the cibriform plate and synapse in the olfactory bulb. The olfactory tract then transmits the olfactory signals to the olfactory cortex at the surface of the temporal lobe. In the limbic system (Fig. 4-3), olfactory information is correlated with feeding behaviour and emotional-motivational behaviour.
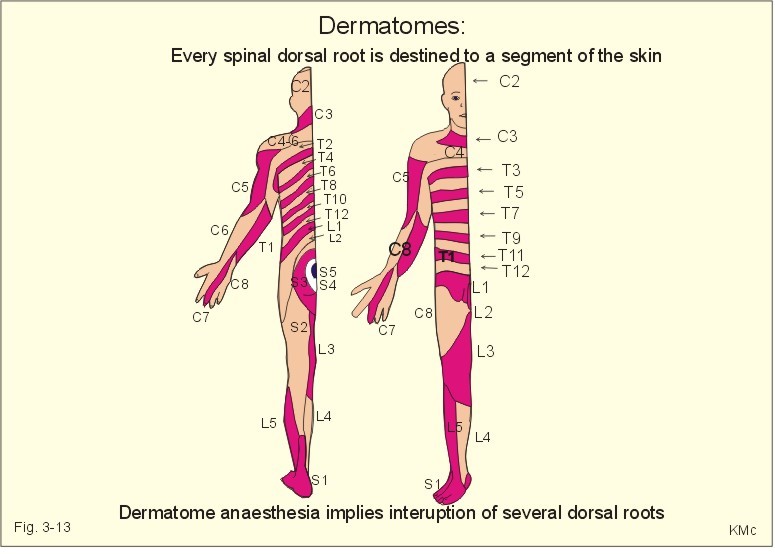
Fig. 3-12: The olfactory region, its receptors and pathways. This paragraph deals with 1.The thalamic syndrome, 2. Brown-Sequards syndrome, 3. Special sensory pain disorders, 4. Taste and smell disorders. The thalamic syndrome is frequently caused by thrombotic blockade of bloodflow to the somatosensory thalamus. The destruction of thalamic neurons in one hemisphere leads to ataxia and loss of sensations from the opposite side of the body. After a few months different types of sensations return, but they are accompanied by pain. 2. The Brown-Sequards syndrome The Brown-Sequards syndrome or paresis includes all effects of transection of only one half of the spinal cord at a certain level. All motor functions on the side of the lesion are blocked in the segments below the level (paresis, spasticity, and loss of vasoconstrictor tone). Sensations of pain and temperature from all lower dermatomes on the opposite side of the body are lost, because of transection of the contralateral spinothalamic tract. The only sensation left on the side of transection is crude touch, because it is transmitted in the opposite ventral spinothalamic tract. The total sensory loss is therefore termed dissociated anaesthesia. Hyperalgesia means hypersensitivity to pain. Hyperalgesia is caused by either hypersensitive pain receptors (sunburned skin), or by facilitated transmission. Facilitated transmission is due to abnormal stimulation of peripheral nerve fibres and neurons of the spinal cord or of the thalamus. A special type of hyperalgesia is present when herpes virus infects one or more dorsal root ganglia. The virus excites the neurons and causes pain in the dermatomal segment subserving the ganglion. The segmental pain circles halfway around the truncus on the affected side. The virus is also transported by axonal flow to the cutaneous terminals, where it causes a characteristic rash confined to the dermatome. The disease is called herpes zoster. Causalgia is hyperalgesia and heat-cold-sensations accompanied by sweat secretion in a region with nerve lesion. The hyperactivity in sympathetic efferent neurons and in nociceptive afferents running along arteries is unexplained. Even the lightest touch at sensitised trigger areas release - within seconds - severe lancinating pain throughout the affected branch of the trigeminal nerve. The cause is unknown, and therapy is usually unsuccessful. Referred pain and central pain Pain that originates from deep organs is poorly localised and often referred coming from superficial structures. This may be explained by the fact that pain signals from viscera are transmitted through neurons in the CNS that also transmit pain signals from a specific area of the skin. Pain due to myocardial ischaemia (angina pectoris) is commonly described as pain originating from the inner side of the left arm, and termed referred pain. The mammalian embryo is segmented into so-called somites, which are innervated by an adjacent part of the spinal cord. Every spinal dorsal root is destined to a segment of the skin called a dermatome. Also muscles (myotomes) bone (sclerotomes), and viscera are related to specific segments of the spinal cord or brain stem. The dermatomes are drawn with sharp borders, which is unrealistic (Fig. 3-13). There is considerable overlap and several successive dorsal roots must be interrupted to produce dermatome anaesthesia. In case of serious spinal cord injury the dermatome map is useful for determination of the level and extent of the lesion.
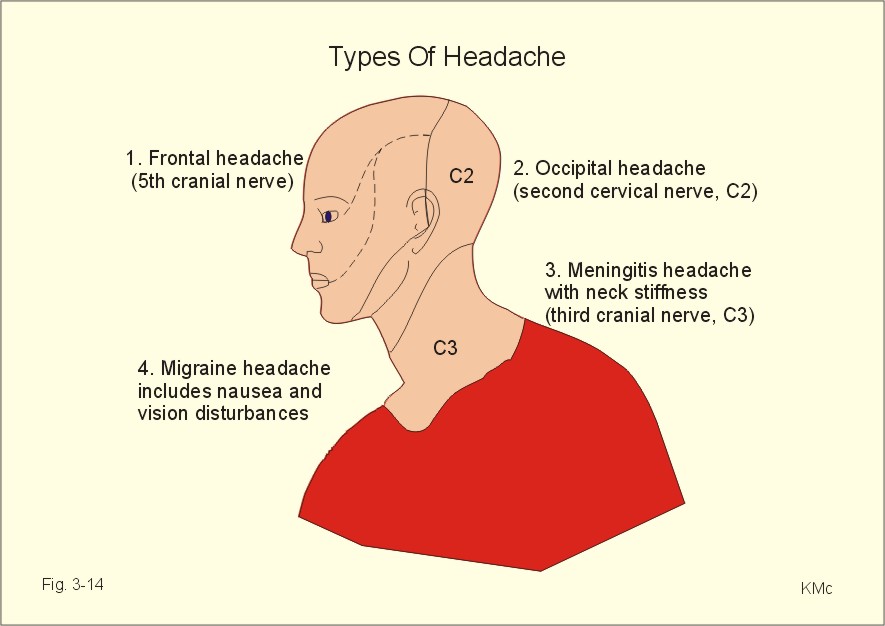
Fig. 3-13: Dermatomes Central pain is a sensation of pain in absence of peripheral nociceptive stimuli. Central pain is processed in the cortical pain areas, and caused by lesions along the nociceptive pathways (peripheral nerves, the spino-thalamo-cortical tract and the thalamus). Amputation of a limb is sometimes followed by phantom limb pain. The patient suffers from severe pains, and the sensation is projected to the amputated limb. It is not known whether the mechanism is central or peripheral. Trigeminal neuralgia (tic douloureux) is a condition with daily paroxysms of violent pain in part of the trigeminal area lasting only some seconds. The pain is provoked by eating, washing the face or by cold in the face. The mandibular and maxillary area is involved. The paroxysm finishes with saliva-tears- and sweat secretion. The disease is probably located to the trigeminal Gasserian ganglion, but the cause is usually unknown. Drugs or surgical procedures on the ganglion have variable effect. Headache is caused by anomalies in intracranial or extracranial structures. 1. Intracranial headache is released from nociceptors in the meninges or in the arteries and veins at the base of the skull. The brain tissue itself hardly contains pain receptors. The intracranial types cover frontal, occipital, migraine, and meningeal psychogenic and pressure headache. Nociceptors are stimulated by stretch (dura or tentorium), by dilatation (vessels), or by chemical means (eg. histamine, 5-hydroxytryptamine etc). The pain signals reach the CNS through the 5th and 9th cranial nerve and the cervical sensory fibres. Stimulation of supratentorial nociceptors is referred to the frontal area via the 5th cranial nerve as frontal headache. Subtentorial nociceptors cause occipital headache through the 2nd cervical nerve. Migraine headache Migraine or hemicrania means unilateral headache, which is frequently but not always present. Migraine is defined as recurrent attacks of headache associated with gastrointestinal and visual disorders. There is evidence for a genetic aetiology of migraine: an autosomal dominant inheritance with reduced penetrance (ie, expression only in permissive environments). Classical migraine has prodromal symptoms (aura) with visual disturbances due to ischaemia in the retina. The onset is often in the eye region with spread towards the vertex or towards the other eye and typically accompanied with nausea, emesis, photophobia and scintillating scotomata. Sometimes sensorimotor abnormalities occur on one side of the body with ataxia, dysphasia and syncope. Frequently migraine occurs without aura, which make the diagnosis difficult. Migraine is of unknown origin, but it is sometimes associated with prolonged psychological stress. A hypothesis claims that prolonged emotional stress in sensitive individuals causes reflex vasoconstriction of intra- and extra-cranial arteries. The brain ischaemia explains the prodromal phenomena, and leads to accumulation of vasodilatating substances such as adenosine, ADP, NO etc. At the onset of the aura, the plasma concentration of 5-hydroxytryptamine rises, and it falls during the migraine attack. After a brief period of aura the vessels dilatate, pulsate forcefully, and the walls become oedematous. These changes are believed to cause the migraine headache. Food containing nitrites and tyramine may precipitate migraine attacks. Severe attacks of migraine are treated with 5-hydroxytryptamine1 agonists, such as sumatriptan, or with ergotamine tartrat. Prophylaxis of migraine is carried out with 5-hydroxytryptamine antagonists (pizotifen, methysergide) and b -adrenergic blockers. Psychogenic headache varies in severity and location. This headache is generally accentuated by conflicts or by anxiety with excessive sweating, tachycardia and hyperreflexia. This type of headache can be the first sign of depression, if the condition is worse in the morning following sleep disturbances. Meningitis headache is accompanied by contraction of the neck muscles (stiff neck). The dura and the venous sinuses are inflamed, and the headache is severe. Pressure headache. Intracranial mass lesions (tumours, abscess, bleeding, and traumata) are usually surrounded by brain oedema, whereby the basal vessels and the meninges are displaced and generate pain. This causes a special pressure headache, which is exaberated by supine rest, bending over, straining, sneezing and coughing. Any elevation of intracranial pressure induces this type of headache. Pressure headache is often accompanied by vomiting. Subdural haematoma must be suspected after head trauma with pressure headache. Suddenly occurring headache following a trauma may be caused by subarachnoid haematoma. When combined with fever, neck stiffness, back stiffness, and vomiting the cause may also be meningitis, where a history of sore throat is frequently obtainable.
Fig. 3-14: The cutaneous innervation of the head and types of headache. 2. Extracranial headache is common and released from nociceptors in extracranial vessels, in the muscles of the head and neck or by inflamed mucous membranes of the sinuses, sinus pain (Fig. 3-14). The pain is felt directly over the frontal or the maxillary sinuses in the case of sinusitis. Typical is the muscle contraction headache located in the frontal or occipital muscles. The frontal and/or occipital-nuchal muscles are tender. Frequently, both the occipital and the cervico-trapezial muscles are tense and tender with specific pain points (loci dolendi). Acupuncture or lazer therapy of these points is often effective. Treatment is difficult when depression or accident sequelae is the underlying cause. The geniculate ganglion is a sensory ganglion for taste, which lies at the genu of the facial nerve. The nerve fibres join the facial nerve in the chorda tympani and carry taste from the anterior two-thirds of the tongue. Cranial lesions involving the petrous temporal bone cause loss of taste in this area together with an unpleasant loud distortion of noise called hyperacusis. Hyperacusis is due to paralysis of the stapedius muscle. The sensory fibres of the glossopharyngeal nerve carry taste from the posterior third of the tongue. Cranial lesions involving the jugular foramen may damage the glossopharyngeal nerve often together with the vagus and the accessory nerve. Loss of the ability to smell is called anosmia. Head injuries involving the cibriform plate or tumours damaging the sensory pathway may cause anosmia. Damage of the olfactory receptors in the nasal mucosa by upper respiratory infections may lead to anosmia.
In his original version, only the exponent n differed for each type of sensation. The equation can be modified by subtracting different constants from SS before raising it to the power n, or by changing the value of k. Chapter 3. Multiple Choice Questions I. Each of the following statements has True/False options: A. The somatosensory thalamus is a relay station for most sensory modalities. B. Glutamate is the main inhibitory transmitter in the CNS, whereas GABA is the dominant excitatory transmitter. C. Pain and temperature reach the thalamus through the lateral spinothalamic tract. D. Presynaptic transmission of opiates inhibits depolarization of nerve cell membranes. E. The adequate stimulus of the cutaneous mechanoreceptors is deformation of the receptor. II. Each of the following statements has False/True options:
A female of 32 years is admitted to a neurosurgical ward with a discrete lesion in the spinal cord caused by a traffic accident. Her vital functions are unaffected. The most important signs are a complete lack of cutaneous temperature sensibility and pain sensibility in the left leg and the lower left side of the trunk below the umbilicus (the navel). Where in her spinal cord is the lesion localised? Try to solve the problems before looking up the answers
Caterina, MJ, MA Schumacher, M Tominaga, TA Rosen, JD Levine, and D Julius. The capsacain receptor: A heat-activated ion channel in the pain pathway. Nature 389: 816-24, 1997. Kandel, E.R., J.M. Schwartz and Jessop. "Principles of neural science." 4th Ed. New York: Elsevier Science Publ. Co., 2000. Russell, M.B. and J. Olesen. "The genetics of migraine without aura and migraine with aura." Cephalalgia 13 (4): 245-8, 1993.
|
||||||||||||||||||||||||||||||||||||||||||||||

.jpg){kind=link}
{kind=link}
Click here to introduce your comments or contributions